
* There was a time when humans didn't know the Earth had a history -- or at least didn't believe it had much of one, being a few thousand years old and fixed in form. That belief crumbled in the 19th century, with the realization that the Earth was immensely old, always undergoing changes in form -- with the organisms that live upon also undergoing changes in form, as elucidated by Charles Darwin. In modern times it is understood, from studies of modern and fossil organisms, that all living entities on Earth are derived from common roots, being arranged in a taxonomic "tree" of relationships.
* Before the era of modern science, the general belief in the West was that the Earth had been around for a few thousand years, with all the creatures created at the start and remaining unchanged to the present day. It wasn't until the beginnings of modern science that this static mentality began to erode. The voyages of discovery conducted by Europeans from the 15th century on led to an explosion of knowledge about the plants and animals of the Earth, and in the 18th century, the Swedish naturalist Carolus Linnaeus (1707:1778) engaged in a massive effort to catalog them all.

In the "Linnaean" system, organisms were organized as distinct "species" or different animals that couldn't interbreed, sometimes with a number of "breeds" or "races" or "varieties" (the last term generally meaning plants) associated with a single species. A number of species could be organized into a "genus" (for example, the genus of cats or "felines"); with a number of genera becoming an "order" (felines, canines, bears, and the like becoming the order of "carnivores"); a number of orders becoming a "class" (carnivores, rodents, primates, and so on becoming the class of "mammals"); and classes becoming members of "kingdoms" (either "animal" or "plant").
Linnaeus was not the first to attempt such a classification, but he was the one who succeeded, his work bolstered by his excellent writing skills, his charismatic ability to influence others, a global network of naturalists feeding him information, and an obsessive thoroughness. The Linnaean classification system, with its scheme of Latin names for organisms (Canis lupus for "wolf", for example), remains in effect to this day, with some additions -- such as subdivisions of the kingdom of animals and plants as "phyla" (the class of mammals being organized with reptiles, birds, fish, and amphibians as the phylum "vertebrates", or creatures with spinal cords). Sometimes plant phyla are referred to as "divisions".
There was little in the system of Linnaeus that contradicted the long-standing notion that species had been created in the past in the same form as they existed in the present -- though late in his life he seems to have had some vague questions about things. During this era of the "Enlightenment", some people were asking more questions about the past history of the Earth. The French naturalist George-Louis Leclerc, Comte de Buffon (1707:1788), presented a new vision of the past history of the Earth in his monumental document NATURAL HISTORY, published in many volumes from 1749 to 1789. Buffon suggested that the Earth and the planets were born from a catastrophe that threw material off from the Sun, with life then arising spontaneously on Earth as the planet cooled.
Buffon also suggested that modern species were derived from different ancestral species. This was clearly proposing the evolution of species, though Buffon was very limited and vague in his proposal, saying that, for example, all the different species of cat were derived from one ancestral species of cat, with no connection between the evolutionary tree of cats and the corresponding tree of wolves and dogs. Other scholars considered the notion of evolution, though none were able to construct a credible case for it.
There was also formidable resistance to the idea. Georges Cuvier (1769:1832) was Buffon's intellectual successor in French naturalism, though absolutely not his heir. Cuvier used his undoubted brilliance and his powerful position in academia to emphatically reject evolution, insisting that as per tradition the species were fixed, having been created and then remaining unchanged to the present. He did admit that there had been changes in species over the history of the Earth. Fossils, the stony remains of the skeletons of ancient organisms, had been long known, but had been dismissed as the remains of freaks or monsters.
Cuvier took them perfectly seriously as evidence of past life -- for example, he examined the skulls of the extinct elephants that would become known as "mammoths" and proclaimed them unarguably different from living elephants. Significantly, Cuvier identified particular types of fossils as associated with particular buried layers of geological deposits, or "strata". He still believed that nature was basically static and unevolving, that the strata were associated with distinct eras that ended in global catastrophes that wiped out species, with a new set of species created in their place. This concept became known as "creationist catastrophism" -- the Maker having implemented all the creatures of each era and then destroyed them after their time was done.
Scientific and public interest in the study of fossilized species -- what is now referred to as "paleontology" -- was high in Cuvier's time. William Smith (1769:1839), a British engineer working for a canal-digging company who learned to pay close attention to what he was digging up, discovered that a particular layer of sedimentary rock could be identified by its distinctive fossils. Smith, as a working person, was not taken very seriously by the British scientific community at first, but other naturalists confirmed the notion.
Cuvier himself was the first to identify a fossil of a great reptile, the first evidence of an age when huge reptiles dominated the Earth. A toothed jawbone well more than a meter long had been dug up from a mine near the river Meuse in the 1770s, but nobody knew what to make of it. In 1795, French troops operating in the region brought it to France, where it was passed on to Cuvier. He identified it as the jawbone of a whale-sized ocean-going monitor lizard -- which was later named Mosasaurus, meaning "Lizard of the Meuse".

This discovery was followed by others. In 1811, a 12-year-old English girl from Dorset named Mary Anning (1799:1847) discovered the fossil of an ocean-going reptile reminiscent of a porpoise, which would be named Icthyosaurus; a fragmentary fossil of this species had actually been discovered in 1699, but nobody knew what to make of it. Anning would become a famous fossil-hunter, discovering another reptilian sea creature, a classic long-necked "sea serpent" that was named Plesiosaurus, in 1821.
William Buckland (1784:1856), a bright, lively, and eccentric don of Corpus Christi College in Oxford, England, found the bones of a big land reptile fossilized in the same strata as Mosasaurus, describing his find in an 1824 paper and calling it Megalosaurus, meaning "Great Lizard". It was a carnivorous beast almost as big as an elephant, that walked on all fours and had a nasty set of sharp teeth. In the meantime, another British naturalist, Gideon Mantell (1790:1852), was picking over the fossil bones of a big herbivorous reptile, which he revealed to the world in an 1825 paper as Iguanodon, meaning "iguana tooth".
Over the following decades, fossil evidence of strange lost worlds of the past began to pile up, with the succession of such worlds increasingly obvious in the fossil record. The geologist Adam Sedgwick (1785:1873), a Cambridge don who, along with William Smith, would be one of the founding fathers of modern geology, probed the earth of Wales and characterized the strata he found. Others were engaged in similar exercises, and in 1841 John Phillips (1800:1874), a nephew of William Smith, published a scheme in which he organized the "eras" of the past as identified by the fossil species observed in the strata.
The oldest strata belonged to what Smith named the "Paleozoic" era, characterized first by invertebrates and then fishes; followed by the "Mesozoic" era, characterized by reptiles, though also yielding early birds and mammals; and finally the "Cenozoic" era, characterized by the predominance of birds and mammals. Each era was divided into "periods", the organization being much like that used today. In 1845, Sedgwick wrote that there was clearly a "progressive development" of forms over eras and periods, but rejected evolution, interpreting the sequence as evidence only of creationist catastrophism.
Other scholars were beginning to question that doctrine -- notably Sir Charles Lyell (1797:1875), another one of geology's founding fathers. Lyell had started out studying to be a lawyer, but he had taken classes from Buckland, a flamboyant lecturer, and got the rock bug. Lyell was inclined towards conservatism about evolution, but in terms of geology he became something of a scientific radical, taking on the catastrophist status quo.
Lyell obtained his inspiration from the work of the Scots scholar James Hutton (1726:1797), who had rejected the idea of a catastrophic history of the Earth, instead proposing that eras of vulcanism -- volcanic activity -- would raise materials from underground to directly or indirectly form islands and continents -- which would then gradually settle into the Earth, to melt and provide another burst of vulcanism. He believed that the Earth lived in a "steady state", with "no trace of a beginning, no prospect of an end." Hutton, in short, had replaced a world a few thousand years old with one that had existed in "deep time".
Hutton's notions were more speculative than rigorously scientific, but Lyell, casting aside catastrophism, was inspired to flesh them out into a much more detailed scientific theory, basing his reasoning on the new geology described by William Smith, Adam Sedgwick, and others, and incorporating the "footprints" of the past as found in fossils. Lyell described his "uniformitarianism" or "gradualism" in the monumental three-volume work PRINCIPLES OF GEOLOGY, published from 1830 into 1833, which envisioned a world of vast time, cyclically reshaped by slow processes of geology.
Hutton had been an all-but-impenetrable writer, but Lyell was crisp and persuasive, bolstered by the instinct established in his legal background to make a clear and persuasive case. Gradualism eventually eclipsed catastrophism -- though many of Lyell's colleagues were reluctant to swallow his insistence that life was basically gradualist as well.
Evolution was finally put on a solid basis by the work of Charles Robert Darwin (1809:1882), a son of an upper-class English family. Darwin was interested in naturalism, having studied under Sedgwick. From 1831 into 1836, he accompanied the Royal Navy brig HMS BEAGLE on a global mapping and survey voyage, in an unpaid position as a naturalist. During the voyage, Darwin investigated the geology and biological diversity of distant lands, coming to the conclusion that the species of the Earth had indeed evolved, all descended from common roots over deep time.
On his return to England, Darwin married and settled down to a life as a gentleman naturalist, spending over two decades fleshing out his evolutionary ideas. It wasn't until 1859 that he decided to go public -- he wouldn't have done it then, except for the fact that he faced being trumped -- with his book ON THE ORIGIN OF SPECIES, one of the most important publications of the 19th century.

Darwin had worked from observations of the succession of species in the geological record; the relationships of species as originally established by Linnaeus; the geographic distribution of species over the Earth; the variation of plants and animals in domestication -- generally through what he called "sports", what we would now call "mutations"; and the equivalent variability of wild species to construct his theory. He had observed how breeders could create new varieties of plants and animals through cross-breeding, using "artificial selection" to work towards a desired variety. Very well; a similar process took place in nature, with simple survival honing variations in species to result, in deep time, in new species, a process he called "natural selection".
In hindsight, Darwin had established a "unified theory" of biology. While there was considerable resistance to his theory of evolution from those who clung to traditional creationism -- there always would be, creationism continues to struggle along in the 21st century -- he obtained adherents in the scientific community immediately. Not all scientists were impressed, however, Darwin himself admitting there was much more to be learned about evolution.
Darwin was particularly weak when it came to the mechanisms of heredity, of which he had the sketchiest ideas, devising an incoherent notion of "pangenesis" as an explanation; even his allies found his thinking on the matter dubious. He did not learn of a research paper, published in 1866 by an Austrian monk named Gregor Mendel that described the patterns of heredity in crossings of pea plants. The paper made little impact on the research community.
The ignorance of the mechanisms of heredity was not really a blow to Darwin's theory; it simply left it without support on that basis, at least until the matter was better understood. More troublesome was the fact that the physicist William Thompson, later Lord Kelvin, suggested from his analysis of the cooling of the Earth that the planet was no more than a few hundred million years old. Darwin admitted that didn't seem like enough "deep time" to permit the evolution of all the species of the Earth; geologists believed Thompson was wrong, but didn't have any way to challenge his figurings.
The result was that by the end of the 19th century, Darwin's theory was in a state of dilapidation. The science community generally accepted evolution; new fossil finds since the publication of THE ORIGIN OF SPECIES had done much to reinforce the idea. However, there was great doubt that Darwin had come close to explaining how it worked. Fortunately, at that same time, new revelations restored life to Darwin's ideas.
The first key to the revival of evolutionary theory was the discovery of radioactivity by the French physicist Anton Henri Becquerel (1852:1908), who found that some materials emitted a mysterious radiation that would fog photographic film. In a few years it would be understood that radioactivity was caused by the energetic breakdown of the atoms of the material themselves, and that the energies produced by such breakdowns were huge -- as given by the famous formulation of the German-American physicist Albert Einstein (1879:1955), "E=MC^2".
The physicists had discovered an energy process that allowed the Universe to be much older than Lord Kelvin had asserted. In addition, since different radioactive materials have different and highly specific decay rates or "half-lives", radioactivity gave geologists a tool to actually estimate the age of many mineral samples. "Deep time" had been restored; not only was the Universe now old enough to allow evolution to take place, it was also possible to provide reasonable hard dates on the fossil record.
The second key rested in the first steps towards the understanding of heredity. It was known at the time that organisms such as plants and animals cells have a central "nucleus"; in the 1880s, microscopic analysis showed that the cell nucleus contained threadlike structures that were eventually named "chromosomes". There were hints that the chromosomes stored hereditary information, but nobody had much idea of the details.
There is no evidence that Gregor Mendel heard of this discovery before his death two years later. Progress forward depended on a Dutch botanist named Hugo de Vries (1848:1935). In the mid-1880s, de Vries began to investigate the breeding of flowers; by that time, it was known that chromosomes were involved in cell replication: they existed in pairs that split up in the germ cells of parents, a process known as "meiosis", and then recombined in the egg cells of progeny.
De Vries and others believed, at the time on somewhat circumstantial evidence, that the chromosomes were carriers of hereditary information. De Vries also suggested, correctly, that different traits were found as "units" positioned on the chromosomes. He had no idea of the precise nature of these "units", but he did give them the name "pangenes", in homage to Darwin's theory of pangenesis. De Vries published his conclusion in 1900. He had resurrected the work of Mendel, bringing it into the mainstream of research. A Danish botanist named Wilhelm Johannsen (1857:1927) gave the then-hypothetical unit of heredity the name "gene" (by stripping down "pangene"), which the British biologist William Bateson (1861:1926) used to mint the word "genetics" for the study of heredity. Later, the entire set of genes for an organism would become known as the "genome".
The first intensive work on chromosome-based genetics was performed by a meticulous American experimentalist, Thomas Hunt Morgan (1866:1945), at Columbia University in New York City, who began work in 1909 on the heredity of the little Drosophila melanogaster fruitfly. They bred very rapidly, and had traits that were easy to observe; Morgan found that they followed much the same general patterns of inheritance as Mendel had observed in pea plants. Morgan deduced that the genes that he presumed existed were strung like beads on a string along a chromosome, and that the recombinations of chromosomes were likely to switch genes around. He was able to map particular genes associated with fruitfly traits on their chromosomes.
Armed with a coarse but workable understanding of the mechanisms of heredity, in the postwar period, biologists were able to establish in the postwar period a much more rigorous model of evolutionary processes, which became known as the "Modern Synthesis" -- which, with its ongoing extensions, constitutes modern evolutionary theory (MET).
Along with a refined theoretical system, biochemists were zeroing in on the precise molecular mechanisms of heredity, discovering that it was encoded in a molecule named "deoxyribonucleic acid (DNA)" that was found in chromosomes. That was puzzling, since DNA didn't seem to be a very diverse molecule. It basically seemed to be a polymeric chain of four "nucleotide" molecules: adenine, thymine, cytosine, and guanine (A, T, C, and G). The challenge touched off a race to determine the actual structure of DNA.
The race was won by a duo at Cambridge University in the UK, including a young American biochemist named James Watson (born 1928), who was investigating DNA in Cambridge, England, working in collaboration with a British physicist named Francis Crick (1916:2004). Working with clues, notably X-ray analysis of DNA crystals, in 1953 Crick and Watson developed their famous "double helix" model of DNA, with DNA actually made up of two linked polymer chains, composed of a pair of linked polymer chains, conceptually resembling a ladder twisted into a spiral. The sequence of the nucleotide "bases" in the DNA molecule clearly defined the genome, providing instructions for the assembly of proteins from their amino-acid building blocks, as well as control functions.
Over the next decades, the understanding of DNA and what it could do emerged into the science of "genomics", in which genomes were decoded and assessed to determine their effects, as well as the relationships of organisms. Genomics became one of the most powerful tools of evolutionary science, allowing the relationships of the Earth's grand family of life, and insights into their past evolutionary history, to be nailed down.
BACK_TO_TOP* The Earth's family of life covers a staggering diversity. Even the job of cataloging it all, referred to as "taxonomy", is a monster job. The cataloging scheme used by modern "taxonomists" was, as mentioned previously, basically invented by Linnaeus in the 18th century. The Linnaen scheme of taxonomy hierarchically broke life down into hierarchical levels of groupings. With some modern extensions, his scheme provides the following "ranks":
Linnaeus did not define the domain and family ranks; they were added later. Modern taxonomists will sometimes insert intermediates such as "suborders" or "superfamilies", and in fact some taxonomists seem to have a fetish for creating new ranks.
Linnaeus devised a distinct naming system for organisms, involving the genus name followed by the species name, both in Latin; by convention the names are in italics, with the genus name starting with an uppercase letter and the species name starting with a lowercase letter. For example, humans are Homo sapiens under the Linnaean scheme, with this name sometimes abbreviated to H. sapiens.
* It must be realized that the Linnaean system is a partly arbitrary ordering scheme, imposed on a nature that isn't structured in such a neat way. This is not to say there's anything fundamentally wrong with Linnaean classification: it's perfectly sensible to create a system to impose order on the complexity of the real world, so we can sort things out. However, it is also sensible to remember that, though there is an order there, taxonomy imposes a structure of labels on it that tends to make it look misleadingly tidier than it really is.
In this document, the assumption is that there are six kingdoms of life. This is just to simplify the discussion, since some taxonomists recognize many more than six, though they are not in any agreement over the exact number. Taxonomists, in fact, have a reputation for being extremely quarrelsome, their style of discussion having been described as "names and nastiness".
Such quarrels can come across as petty, but there is some basis for them. Classification of anything may be difficult -- it can be troublesome to figure out a consistent scheme of organizing a large collection of photographs or the like. To be sure, the branching tree of life means that the familial relationships among organisms are not at all arbitrary: of all conceivable sets of relationships, only one can really be right. The problem is figuring out exactly which among the possible relationships are the right ones. Clearly unrelated species can sometimes look surprisingly alike -- sharks and dolphins are a good example, though taxonomists haven't had any trouble recognizing them as unrelated for a long time. On the flip side, sometimes related species can look very different -- whales and hippos being the prime example, most people being very surprised that the two are closely related.
There's also the difficulty of distinguishing closely related organisms as separate species: dogs and cats are unarguably separate species, they couldn't interbreed if they tried, but what about cats and wildcats? Some types of wildcats clearly resemble housecats; are they a separate species? What about more distinctive types of wildcats, such as lynxes and caracals? In borderline cases, that is a matter of opinion. The notion of a "species" is misleading in that it makes it easy to think in terms of a "model number" of a machine, a "Mark I" or "Mark II" or whatever. Models of machines are distinct and well-defined; not so for closely related organisms.

* In the beginning of proper taxonomy, the only real tool taxonomists had was homologies, the resemblances between different species of external and in particular internal biostructures. That was workable to an extent, but homologies could also prove misleading, as noted above. In Darwin's time, the analysis of embryos provided another useful tool, though again one that could be misleading. Determining the correct taxonomic relationship between species requires the integration of multiple aspects of organisms -- and the more aspects, the better.
In modern times, analysis of protein structures has provided an excellent tool for understanding relationships between species. One of the most popular proteins for taxonomic work is "cytochrome c", a component of the cellular energy production system in animals, plants, and fungi that consists of about a hundred amino acids. It is useful because it covers such a range of species. Assessing the similarities and differences between the cytochrome c proteins of different species gives an index of their relatedness.
A variety of genetic analysis tools are also now available -- they are discussed later -- that have given taxonomists a big leg up, handed them a few big surprises, and resolved a good number of long-standing disputes. They still bicker, but it's generally over subtler distinctions. Experts can be defined as people who argue more and more over less and less, until they're arguing all the time over nothing at all.
BACK_TO_TOP* Modern taxonomy is based on the concept of the "clade". The notion was introduced by the German entomologist Willi Hennig (1913:1976) in the 1950s, though it didn't really hit the scientific mainstream until the 1970s.
A clade is a strict form of family grouping, defining a hierarchy of a particular type and all of its variants. This looks very much like traditional taxonomy in some respects, but it differs in others. For example, in traditional taxonomy reptiles and birds were separate classes, but under cladistics, classification has become more troublesome. As defined under traditional taxonomy, reptiles were not a clade, since birds are derived from them, meaning some of the family members had been excluded. By the old definition, reptiles were "paraphyletic", meaning they didn't make up a complete "monophyletic" clade. The definition of reptiles has been considerably adjusted in recent times to conform to cladistic requirements, a matter also discussed in more and somewhat confusing detail later.
Sometimes such paraphyletic groupings are called "grades" instead of clades -- if just for convenience, since it's identifying a "family" with a large chunk of the actual members thrown out. In the vaguest case, a grade is nothing more than a "polyphyletic" grab-bag of organisms with some specific features in common and with no strong relationships between them. That's why Hennig invented "cladistics" in the first place, to ensure that taxonomists recognized true familial relationships, and weren't just cataloging species by some arbitrary and possibly prejudiced scheme of convenience that didn't necessarily reflect the actual relationships. There's nothing wrong with convenience, of course, and grades are still used in taxonomic literature, though there is a push to designate them by attaching a "*" to the name.
Clades also do not by themselves define ranks, levels of hierarchy, as do traditional Linnaean taxonomic designations. Mammals by themselves form a clade; carnivores form a subordinate clade; the various cat species form a clade beneath that. In addition, clade diagrams or "cladograms" are not "evolutionary trees" as such: they provide a branching tree terminating in offshoots, living and extinct, along branches of variations that don't feature any species themselves, though they may have labels giving the name of the clade defined by particular branches.
In fact, to the strict, clades are only defined by taxonomic features, not by supposed ancestry -- the "variants" mentioned above are only variants in the sense of the relationship of their taxonomic features. Taxonomically, chimps and humans are more closely related to each other than they are to any other animals, but that fact is established by their taxonomic features, and remains a fact whether anybody buys evolution or not. Some taxonomists even go so far as to declare evolutionary science "irrelevant", but that's just a provocative way of saying they don't think it's part of their particular job.
Since features are a product of ancestry, cladograms do strongly follow evolutionary patterns, and they would make little sense without assuming common descent, but the diagrams are constructed on the basis of taxonomic features that imply paths of descent -- not the other way around. Hennig's goal was just to reduce the clutter that made taxonomy more difficult, and sometimes led taxonomists to classify organisms on the basis of assumed ancestral relationships that weren't supported by taxonomic features and sometimes didn't exist.
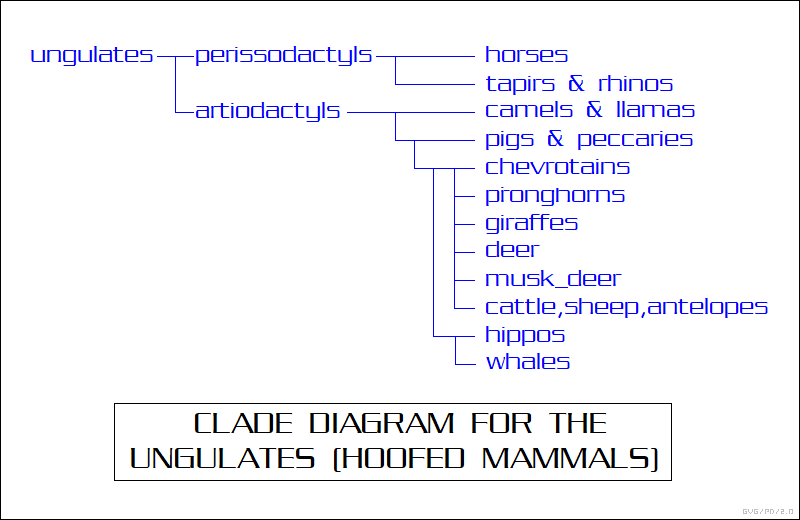
In its early days, cladistics introduced an entirely new level of dispute to the contentious field of taxonomy, though it is generally accepted now. However, these disputes were really not of much interest to anyone but taxonomists, and there was never much objection among them to the idea that this enormous array of organisms evolved by natural selection over time, branching out in ever-increasing elaboration from the origins of life billions of years ago.
One of the implications of cladistics is a clash with traditional Linnaean taxonomy, and in fact some taxonomists have called for junking the Linnaean system. The general consensus for the time being is that there is a trade-off between the rigor of cladistics and the convenience of Linnaean classification. We lump the animals we call "reptiles" together because they have clear resemblances: they have scaly skin, they are cold-blooded, they lay eggs. As long as we realize that birds are descendants of reptiles, this causes no confusion; and as it has been pointed out, any researcher actually working in a particular domain of biology knows what's going on and finds the Linnaen classifications to be no particular bother. The Linnaean scheme is too useful and convenient to simply discard, but it should be treated flexibly, being extended or modified as needed.
BACK_TO_TOP* Pinning down the emergence of the tree of life is not easy. The main evidence is from fossilized organisms dug up from the earth. The first difficulty is that homologies are the only taxonomic tool available for investigation of long-extinct species. Homologies are workable, but they are not in a league with proteins and DNA -- which are, as a rule, only available for species that went extinct recently, and the older the samples, the more difficult they are to examine. That makes it even harder to sort out species of fossil organisms than of living ones. In Victorian times fossil-hunters were careless, quick to identify any odd bone or skullcap as belonging to a new species in something of an exercise of racking up scores, but modern paleontologists are much more thorough and cautious.
Finding fossils is troublesome. Fossilization is an unusual circumstance: after most organisms die, they are devoured or decay, and little trace of them is left. Fossilization generally requires that organisms be neatly buried in sediments, ash, or the like, and left undisturbed while their environment turns into stone. Shallow-water creatures also tend to be disproportionately represented in the fossil record, because the bottom sediments of such regions are the most optimum environment for creating fossils. Soft-bodied organisms tend to be poor candidates for fossils, and so creatures with shells tend to be disproportionately represented in the fossil record. Big creatures tend to be more easily preserved than small ones -- though small ones like insects do end up being trapped in plant pitch, to be unusually well preserved in lumps of amber. Examination of amber samples has become a boom business as of late, with paleontologists able to performed detailed examinations of organisms in opaque amber using X-ray tomography technology.

In addition, it's not likely to find fossils by digging at random; they are generally found in isolated fossil beds. Worse, even stone is impermanent, with geological processes tending to disturb or destroy fossil beds over time, meaning that the older the fossils, the scarcer they tend to be. In fact, it's almost impossible to find undisturbed rocks over about 3.5 billion years old.
Even when fossils are found, they're rarely pristine and detailed, and are also often incomplete -- with some paleontologists describing their most prized fossil finds as "road kills". In fact, paleontologists have a bad reputation for placing too much weight on organisms only known from their teeth, some suggesting that a field of study named "toothology" needs to be set up. To sure, teeth are relatively easily preserved, and they are informative remains; if we find a giant shark's tooth, it's persuasive evidence of a giant shark. Extrapolating the properties of an organism from teeth that are less familiar can be dodgy.
* Fortunately, there's no shortage of much more detailed fossils, and absolutely no shortage of fossils. There have been vast numbers of organisms on this planet, and even if only one in millions survives as a fossil, that still gives plenty of clues. In addition, the fact that geological layers are consecutive, carry signatures in the form of distinctive fossils, and can be dated by radioactive methods allows a chronology to be assigned to these fossil clues. The dating methods allow the relative ages of geologic layers to be pinned down, even when the earth has been flipped over by folding processes, putting the oldest layers on top.

The fossil record, for all its defects, has created a generally consistent vision of just how life has evolved on Earth. Even a sketchy record provides plenty of information. Suppose we take a video, for example an old Charlie Chaplin silent movie, and get a set of consecutively-numbered prints selected at random from about one in every hundred frames -- which is about, on the average, one print per every four seconds of video. That would still be enough to get a general idea of the evolution of the movie's story, though there would be mysteries here and there. Of course, that's assuming that the movie features a coherent, continuously developed narrative. If a movie cut down in this fashion were loaded with flashbacks or completely incoherent, we'd get disjoint images; we'd be hard-pressed to figure out what was going on, and would realize we didn't have a story that made sense.
Similarly, suppose we only know about one in a hundred of the clearly distinct large-scale organisms that have ever existed on the Earth. We still have enough information to get a general idea of the story, and to see that there is indeed a coherent evolutionary story and not just a set of disjoint images of the past. Despite the fact that there are mysteries here and there -- which would remain mysteries no matter whether MET was correct or not -- the overall scheme remains visible. The successive emergence of different types of organisms is very clear, and is not violated.
While the fossil history may be incomplete for particular species, the evolutionary tree becomes increasingly obvious as the taxonomic level rises, and as discussed later there are more than a few specific cases in which the evolutionary evidence at the detail level is actually very impressive. In addition, as new fossils have been discovered, they have generally fleshed out the tree, requiring no more than adjustments in details of the story. Given the current level of activity in fossil-hunting and the availability of good sites, paleontologists believe they are now entering a period where new fossils will be discovered at a rate much greater than in the past -- and would take odds that these new discoveries will also flesh out the story as it is known, and not turn it on its head.
In sum, as far as MET is concerned, the fossil record is not at all a problem, in fact it's all for the good. Nobody who's ever visited even a modest museum of natural history would find its fossil collection a joke; all that has been discovered so far buttresses MET, and the discoveries keep on coming in at an ever-faster rate. The reality is that the fossil record is much more generous than we might expect to be.
BACK_TO_TOP
