
* Much of the evidence for evolution was outlined by Darwin himself, and remains just as valid now as it was in 1859. This chapter and the following recapitulates and updates some of his core arguments.
* Darwin partially based his theory on consideration of the taxonomic tree of species originally laid out by Linnaeus, and the tree of life is still regarded as a cornerstone of the concept. The reasoning starts out as a bit subtle and arguable, but builds up credibility as it is developed.
The tree of life clearly shows the existence of families of organisms. Among the mammalian carnivores there is a family of cats, such as lions, tigers, cheetahs, and wildcats; a family of dogs, such as wolves, jackals, foxes, and domestic dogs; a family of bears, including grizzly bears, sun bears, and panda bears; and so on. The reality of this familial tree was well established, on the basis of physical details, even as far back as Darwin's time, though there was considerable dispute over special cases and the details. As discussed later, modern genetic evidence has confirmed that the taxonomists of the past were generally on the right track, though many tweaky corrections and elaborations have been introduced.
The idea that there is a range of species collected in different families is not on the face of it incompatible with creationism, since it can be argued that different creatures were Designed for different roles and that a set of familial templates or a set of "common Design" principles were involved in the "Intelligent Design" process. As mentioned, Richard Owen presented this argument in response to Darwin early on, and it is still floating around today.
In reality, the entire concept of "biological relationships" strongly implies a "familial relationship", and the notion of a family implies common descent. If there were some isolated human community where the population could be roughly arranged in three groups with common names and distinct physical similarities within each group, it wouldn't be a shock to think that the entire population of the community descended from three distinctively different founder families. It's just as natural to think that all the different species of cats had a common ancestor.
The basic problem with the "common Design" argument is that it really doesn't refute the notion of evolutionary descent of organisms -- because it invokes the analogy of common design in machinery, which is clearly due to evolutionary descent of machines. Human designs evolve; elements introduced in one generation of a machine are reused in its descendants. It may be straightforward to devise an evolutionary tree for a family of products, such as a series of aircraft. Lines of progress in human invention tend to resemble biological evolution because humans necessarily build on what they already know; it is humans, in effect, who are imitating nature, which hardly implies that nature imitates humans.
* The tree of life was an important element in Darwin's case, and it was closely related to the notion now referred to as "speciation" -- the division of a single ancestral stock into or more species. This is a subtle concept, with considerable debate over the details even today. One subtlety that Darwin understood perfectly well was the interesting question of why there are more or less distinct species in the first place.
If, say, all the different species of antelopes evolved from a common ancestral species, why do we see relatively distinct species of antelopes? Why not a continuous spectrum of different "flavors" of antelopes? Why do we have recognizable antelope species like kudus, duikers, impalas, gerenuks? What Darwin was basically asking was: why species? Or stated somewhat differently: What are species?
The traditional definition of a species, as mentioned earlier, is of a population of interbreeding, or at least potentially interbreeding, organisms. A species is not like a particular production model of a machine, with all units produced being effectively identical. Humans vary over a wide range of height, weight, skin color, hair color, but they still feature a large degree of commonality in form that would make them clearly identifiable as a single species to any aliens who decided to pay our planet a visit.
The variation in features among our species could be represented by the "bell-shaped curve" often used in statistical analysis -- at least in an abstract way, since the range of different features is vast, and they may not follow a nice smooth curve. Such an abstract curve would have a median (central) value of the "average" human, being of average height, weight, and so on, and could be visualized as representing the "less fit" to the left of the median and the "more fit" to the right.
Any sexually functional male human can in principle reproduce with any sexually functional female human. Sexual exchanges of genes within the human population establish the shared "gene pool" that is responsible for the range of traits of humans. Similarly, a population of a particular species of antelope, say gerenuks, consists of individuals who can successfully reproduce among each other and who share a range of traits, with a gene pool corresponding to those traits. That gene pool is not static -- it will change over generations as mutations arise in individual antelopes and are added to the gene pool, while selection gradually winnows out individuals at the left end of the abstract bell curve, shifting the curve to the right. For example, since gerenuks are relatively small antelopes, it's a fair if not certain bet that members of their ancestral population were bigger, but over time slowly shifted for whatever selective reasons to a smaller average size.

Suppose a population of one species of antelope is split, say by the shift in course of a river that is too big for the antelopes to get across. Now there are two populations that cannot crossbreed, or in other words they are "reproductively isolated". Initially both have more or less the same gene pool, but over time as mutations arise and selection has its effect, the two populations are going to become ever more different through a process of "drift", each acquiring different features.
The interesting thing is what happens if the river shifts back and the two populations of antelopes come in contact again. If their reproductive isolation was only for a relatively short time, nothing too exciting happens, they start sharing genes again and create a gene pool merged from both populations. The end result is a population of hybrids -- which, incidentally, may be distinctly different in features from either of the two populations it was derived from. However, if the isolation lasted for a long time, the two populations may have become too genetically different to breed together well or at all. They have become two species. Even if they did remain genetically compatible after their isolation, they may have acquired features, such as different sex colorations, that keep the two populations from sharing genes; they will continue to drift until they can't interbreed even if they were inclined to.
An interbreeding population tends to maintain a common gene pool, while reproductive isolation tends to split a population along down different paths that gradually set up barriers to re-consolidation of the gene pool. Over some fair interval of time, two new species emerge. The process of speciation is why there are distinct species -- and not a continuous range of descendants from a common ancestor.
Incidentally, species may arise without a geographic split in the population, for example through the emergence of differences in sexual behavior -- female preferences for coloration for example -- in one population that gradually split it into two non-interbreeding populations, even though the two populations were always mingled with each other. Such "allotropic" or "sympatric" speciation might be expected to be rare, but it appears to depend somewhat on the sort of organisms involved. It's certainly rare for, say, mammals, but it appears it may be fairly common in others such as marine plankton of various sorts. It may be reducible to an issue in system dynamics, that some changes will tend to be absorbed back into a population of organisms, while others will create an instability in the genomic makeup of the population that, due to the introduction of a reproductive bias, is amplified until it splits the population in two.
* The process of speciation is clearly illustrated by the fact that the boundaries between members of a species are often indistinct. Wolves can interbreed with dogs -- in practice usually the larger ones, because wolves tend to regard smaller dogs as prey -- and produce fully functional offspring. Lions and tigers can produce offspring, but they are generally sterile. More widely differing species cannot interbreed at all.
Wolves and dogs are still close together on the taxonomic tree, implying they only split apart recently. Lions and tigers are farther apart, which suggests they split apart some longer time ago. Interestingly, about ten percent of human couples can't conceive, and of these couples roughly a tenth are perfectly fit as far as their reproductive facilities go; the only explanation anyone has come up with is that they are suffering from a genetic incompatibility. To be sure, nobody disputes that all humans are the same species, but the couple may well have specific features in their genomes that make it difficult for them to pair up. The couple in effect came from the opposite ends of a "genetic compatibility" distribution; had the husband and wife chosen different mates closer to the center of the distribution, neither would have had any problems.
If this is so, it would be strong evidence for the way that gradual genetic drift can set up reproductive barriers. There are clearer examples of the emergence of such reproductive barriers in nature, for instance with pink salmon. Since salmon have a life-cycle that takes them up rivers and streams to spawn every two years, that means there are two groups of salmon, "even-year" salmon and "odd-year" salmon, that don't normally breed with each other. As it turns out, it's difficult to cross-breed even-year and odd-year salmon, despite the fact that they are hard to tell apart.
* It was the process of speciation that produced the species known today from common ancestors, with the patterns of division reflected by the tree of life. The tree of life makes very little sense without assuming common descent; up to the era of Linnaeus and Darwin there were a large number of theories put forth to explain the taxonomic organization of species, but in hindsight they sound like little more than arbitrary schemes for categorizing the books in a library. Much the same could be said for a modern taxonomic scheme proposed by creationists sometimes known as "baraminology", in which organisms are divided into groups of "created kinds" -- that is, a handful of basic "kinds" were "Intelligently Designed", with microevolution then leading to variations within (but not outside of) each "kind".
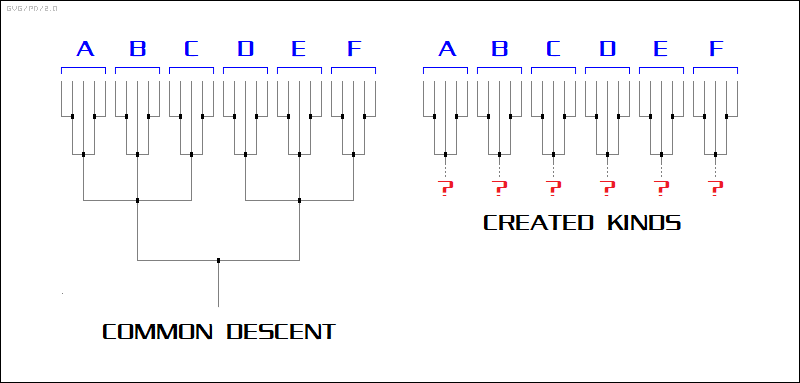
The notion of "created kinds" is essentially a variation on the game of microevolution versus macroevolution. It concedes the branches of the tree but then, proclaiming some form of "magic barrier", declares the trunk underneath nonexistent. Creationists also like to claim that the failure to find bizarre hybrid organisms -- the most famous being the "crocodile-duck" or "crocoduck", with the head of a crocodile and the body of a duck -- as demonstrating that evolution is wrong. Actually, such a contraption would make absolutely no sense in the context of MET, or for that matter in any other context outside of P.T. Barnum.
One could just as easily claim that if all dogs are actually descended from wolves -- and they are, no sensible argument about it -- then we would have to expect the existence of a "great chihuahua", with the big body of a great dane and the little head of a chihuahua, or for that matter the reverse. One could imagine a wide range of such chimeric dogs, but it would have to be for fun, since the simple fact of the matter is that the ancestor of all dogs looked like a wolf.
BACK_TO_TOP* Now consider the way species vary with location. All species are subject to a greater or lesser degree of genetic drift all the time, obtaining genetic changes that either have no visible effect or result in traits even fairly significant ones, that neither impair nor enhance survivability. Under MET, separated members of the same species inevitably become increasingly dissimilar.
Why, for a prominent example, are the organisms of Africa and South America so different? Why are there pangolins -- scaly anteaters -- in Africa and armadillos in South America? Why are there aardvarks in Africa and anteaters in South America? In both these comparisons the animals are clearly not closely related despite their significant similarities. Most significantly, there are distinctive families of primates in the Old World and New World, with easily identified distinctions. There are no baboons in the New World; there are no marmosets in the Old World. This sort of geographical distribution of families of organisms fascinated Darwin, and he appropriately made it a major element of his theory.
The biogeographical evidence for branching evolution becomes more evident when regions that were long ago joined to large land masses are disconnected by continental drift or some other mechanism. The large island of Madagascar is noted for its unusual species. Madagascar has been geologically isolated for 90 million years; although it is not all that far from Africa in the current era, its wildlife is almost entirely different from that of Africa, in particular lacking any large native species of ungulates. It includes a fascinating set of species of lemurs, which are only represented in Africa by the bush babies and pottos, while there are no monkeys or apes in Madagascar.
It appears that the ancestors of Madagascar's lemurs arrived tens of millions of years ago by a "rafting" event, in which they floated over the sea on uprooted trees or the like. This sounds like an implausible notion, but it has been observed in modern times, and in fact hurricanes can tear up so much vegetation that the rafts look like small islands, swarming with various animals. It is admittedly rare -- but over millions of years, events of low probability become a near certainty.
In any case, since that time, their lineage has been pushed to one side in Africa by the monkeys and apes, while they thrived in Madagascar. There was once a huge ground-living lemur there, the size of a bear, and in the current day the family includes, in sadly small and shrinking numbers, the notoriously bizarre nocturnal "aye-aye", named for its cry. It is the ecological equivalent of a woodpecker, with forepaws featuring a long and thin middle finger, which it uses to search for and dig out insects burrowed into trees. Australia is an even more impressive example, with its large numbers of species very different from those found elsewhere -- in particular its large number of "marsupial" mammals -- that is, mammals that rear their young after a short gestation in a pouch, as opposed to the "placental" mammals, in which young are born much more fully developed.
* Where the evidence becomes particularly impressive is with isolated and remote island groups that had never been associated with a large landmass and are inaccessible to organisms that can't cross large stretches of salt water. Darwin keenly recognized the importance of such cases, with the distribution of finches among the separate islands of the Galapagos ending up being a particularly interesting example. The pattern of the species of Darwin's finches suggested that ancestral finches arrived from South America and became established, radiating out into species differing in appearance and habit.
The only organisms that can reach these islands are those that can fly to them, swim to them, or hitch a ride somehow to them on things that can fly, swim, or float. That's why such isolated islands rarely featured mammals aside from bats or seals, at least before the arrival of humans and their camp followers. Reptiles were also uncommon -- the Galapagos are unusual in featuring significant numbers of native reptiles, but they are all clearly variants of a single ancestral iguana stock.
Such isolated island had birds, of course, and insects that floated in on the breeze; plants were restricted to those whose seeds could be blown in on the winds, float over the sea water, or be carried on the toes and feathers of birds. When the islands were clustered in chains, as in the Galapagos, the results were particularly striking, with different variants of organisms derived from common stock on different islands. The Hawaiian Islands demonstrate a similar pattern of species, particularly the group of birds known as "honeycreepers" -- which diversified in their isolation into a much wider range of variation than the Galapagos Finches.
Such isolated species as the honeycreepers, existing in environments where they aren't confronted by many threats, tend to be very vulnerable to invasive species, like cats, rats, and pigs brought by humans. Most species of Hawaiian honeycreepers are extinct, and there are real worries that those that remain won't be around much longer.
* One of the more spectacular illustrations of speciation concerns the "cichlid" (pronounced "sick-lid") fish of Lake Victoria in Africa. Cichlids are bony fish with an unusual adaptation: they have a second set of jaws set back in the throat. That would be intriguing in itself, but what makes the cichlids of Lake Victoria more fascinating is that there are hundreds of species of them, all closely related to each other, and relatively distantly related to other cichlids. There are also cichlids in Lake Tanganyika and Lake Malawi, but the species are different in each of the three lakes.

Lake Victoria was formed about 100,000 years ago, setting a time bound on how long it took to produce the hundreds of species of cichlids in the lake. In addition, borings from the bottom of Lake Victoria show that is carpeted with a layer of dead grass, dated from 14,500 years ago. At that time it largely dried up, being reduced to a set of separate small lakes and ponds. Each pond became a separate ecosystem, an "inverted island" of water surrounded by land, in which the cichlids underwent rapid diversification. Once the lake filled back in again, the expanded numbers of species became parts of an expanded ecosystem. It appears that this cycle occurred several times over the last 100,000 years, driving the wild speciation of the cichlids.
Incidentally, there is the peculiar case of two small lakes in the Cameroons that each have a number of different cichlid species -- one lake with 9 species, another with 11. In both cases, the cichlids of the lake are clearly derived from a common ancestor, forming a separate branch of the cichlid family tree from cichlids in other lakes, meaning that they arose in the lakes and did not arrive from elsewhere. The peculiarity of the matter is that these lakes are too small to have supported any isolation of a single population into multiple groups, separating their gene pools so they could have evolved into different species. It appears to have been sympatric speciation at work, though the specifics are unclear.
* There are anomalies in geographic distribution that were once puzzling, but turned out to make perfect sense. For example, the big ratite birds are found in very distantly separated regions -- the ostrich in Africa, the emu in Australia, the rhea in South America. It wasn't until the discovery of continental drift that this pattern of distribution was understood. The ratites are an ancient lineage by bird standards, and at the time of their origin these continents were linked: they simply walked from continent to continent, and were then isolated when the continents drew apart.
Sometimes such apparent anomalies end up confirming the rule instead of breaking it. Camels are found in the Old World, while their close relative the llama and its kin are found in South America. This was puzzling to researchers at first, leading to the conjecture that camels were once common in North America but died out. Fossils of extinct camels were eventually discovered in North America, including those of the giraffe-like Aepycamelus AKA Alticamelus. In fact, it seems that camels originally came from North America. There were times when the Americas and Eurasia were connected, allowing organisms to cross both ways. Camels were among those that went west; cats were among those that went east. This is why it isn't surprising that genetic analysis shows jaguars to be close relatives of tigers and lions.
Similarly, marsupials are not at all found in Africa and Eurasia even as fossils, but they are found in both Australia and South America. That was puzzling before the discovery of continental drift, but it is now known that the two continents were joined together in the timeframe of the origins of marsupials. They were both joined to Antarctica, which was not as frigid in the distant past as it is now -- and marsupial fossils have been found in Antarctica.
BACK_TO_TOP* The discussion of the evolution of the eye demonstrated a basic principle of evolution: the spectrum of forms of a particular structure, illustrating how the structure could have emerged in an evolutionary fashion. MET insists that species emerge by slow microevolutionary changes, one step at a time -- "natura non facit saltum". Furthermore, as implied in the discussion of the evolution of the eye, each one of these changes should provide a selective advantage over the status quo, or at least do no harm. It has been said that evolution is like an architect who can never design a completely new building -- the architect can only design a minor variation on a previous one, with every building in the sequence as or more habitable than the previous one.
In more formal terms, evolution requires that there will be no "non-functional intermediates" in an evolutionary sequence. The notion of non-functional intermediates seems to raise a challenge to MET. It's like the story about Alice trying to get across a canyon to visit Bob, rearranged to have Alice get across a river by walking on sets of stepping stones that were luckily arranged to allow her to make the crossing one sure, solid step at a time. What are the odds? They don't sound good, and they sound like they should get worse for greater evolutionary changes, since that would mean more lucky stones.
This is a made-up problem. Consider a scenario for the evolution of a particular organism from an earlier species, visualized again as a video with one frame per generation to show the gradual transition of one species into another. The video seems to show the organism changing smoothly and progressively in a magically directed fashion from beginning to end. It only looks like that because the video was constructed with a particular endpoint as an anchor: it is viewed in the forward direction, but it was assembled by retracing the steps from the endpoint back to its origin.
The organism is actually evolving in a range of different directions per generation. Several videos could be made to show the same organism evolve into distinctively different species -- think of Darwin's finches -- though most of the branches are dead ends, often dying out quickly because the modification didn't provide any selective advantage, or immediately because it was downright disastrous.
The story also assumes that Alice is trying to find her way to Bob, but that isn't how things work. Another homely model that can be used as an analogy for evolutionary processes is to imagine a robot car that cruises the US interstate highways, taking interchanges between highways at random. The idea that such a dumb robot could navigate from New York City across the continent to Los Angeles seems preposterous, and in fact it is. However, barring accidents, the car is going to end up someplace: maybe Dallas, maybe Miami, maybe Toronto, maybe Seattle, maybe Chicago, maybe even Los Angeles. Similarly, Alice is simply walking wherever she can put her foot down and has no thought of where in particular she is going; she's not trying to get to Bob's house, it's just where she happened to end up. If she couldn't take a particular step, she just didn't go that way. Claiming a problem is invoking the "prosecutor's fallacy" again.
Creationists often like to perform analyses of evolution's deficiencies as a "search algorithm" -- usually coupled to more traditional "monkeys & typewriters" dubious probability calculations -- to show that it can't possibly work. However, thinking in terms of search algorithms is getting off on the wrong foot at the outset, since evolution isn't searching for anything; it's not going anywhere in particular, except in the direction of improved fitness.
Evolution isn't a search algorithm, it's a sorting algorithm; discarding the less fit at a greater rate than the more fit. In any generation of an organism, there's likely to be tiny changes among some of the members of the population that provide a slight selective advantage -- any changes, any selective advantage. Again, the fitness landscape, while it is visualized in terms of hills and valleys, is a sheer abstraction of a "surface" with many dimensions, with every applicable measure of "fitness" adding its own dimension. That by implication means that there's a lot of possible "uphill" paths -- if the organism reaches a peak by one measure of fitness, it can still be moving uphill by some other measure. Neutral variations also play a big role, with the evobot drifting haphazardly over the fitness landscape in regions where it is flat.
The question of non-functional intermediates seems valid at first, but it ends up being a red herring, another attempt to declare a "magic barrier". Once again, the language analogy holds. One language will transition into another over time through slow, small, unplanned changes. None of the changes are going to take hold unless the speakers of the language find them useful, advantageous. At all steps, the language remains fully functional -- there's never any unworkable "intermediate language". The process could proceed until there is no recognizable resemblance between the two languages and never run into any "magic barrier", and there is no "blueprint" to show where it will end up: languages are not, as a rule, the product of a predetermined overall design.
* Anyway, back to the example of the eye. It can be shown to have evolved by tiny steps, each of which gave a selective advantage. What good is 5% of an eye? A primitive eyespot isn't even that much of an eye, but it's still useful. By the same reasoning, consider the "sea dragon", a species of seahorse that looks exactly like a clump of seaweed. Would it do any good to look only 5% like a clump of seaweed? Yes -- wouldn't a fish that looked 5% like seaweed have something like a 5% advantage in camouflage? Wouldn't a predator have more trouble spotting it from a distance or under poor viewing conditions? Evolution is based on such incremental advantages.

It should be emphasized again that natural selection works on populations; "a fish" doesn't mean a single fish, it means a population of interbreeding fish. That population will have a variation in characteristics around a mean value of something like a bell-shaped curve. It's not really a simple question of two fishes -- one that has achieved a jump in its resemblance to seaweed relative to the other -- but of an entire population in which there is a continuous range of appearances, with one end of the range clearly looking less like seaweed and the other end of the range more like seaweed.
The fish on the outlier of that curve that looked less like seaweed would be slightly more vulnerable to predators than those on the other outlier of the curve that looked more like seaweed, causing the mean of the curve to very gradually shift towards the component of the population that looked more like seaweed. This cycle of ever improving seaweed impersonations started by accident: due to its ancestral history and other adaptations, the fish already looked slightly like seaweed, and random changes then enhanced the effect, with natural selection ramping up the evolutionary treadmill through its "differential screening".
Eventually the fish obtains a highly optimum resemblance to seaweed, and the probability of obtaining a mutation that improves the resemblance starts falling off, while the probability increases of mutations that will degrade it. Incidentally, there are two sorts of sea dragons, "leafy" and "reedy", that impersonate different sorts of seaweed. In any case, the fish has effectively reached the peak of that hill on the fitness landscape, and then the differential screening will trim away at both ends of the distribution, squeezing it in towards the center and resulting in a highly uniform, well-tuned population of fish that look surprisingly like seaweed. Selection pressures are no longer promoting change; instead, they are promoting stability.
It should be remembered that the fish undergoing spontaneous genetic changes that take it off in many evolutionary directions in parallel. Some of the changes will reduce its fitness, and be dropped out of the population gene pool sooner or later; others will be beneficial, and will be retained. While the fish is gradually adapting to a camouflaged form, it may also be acquiring refinements of its digestive processes to suit it for its particular diet. Of course, this is not a neat or optimized process; changes that improve its advantage in some respects may end up being at the expense of a disadvantage in other respects, for example a loss of some of its neat resemblance to seaweed. Two steps forward, one step back; it may not seem sensible, but that's the way it works.
For another example, consider aquatic adaptations. The paleontologist Stephen Jay Gould, well-known for his popular science essays, once quoted a creationist who suggested that adaptation of mammals to a marine environment posed a major problem for MET:
QUOTE:
Darwinists [sic] rarely mention the whale because it presents them with one of their most insoluble problems. They believe that somehow a whale must have evolved from an ordinary land-dwelling animal, which took to the sea and lost its legs ... A land mammal that was in the process of becoming a whale would fall between two stools -- it would not be fitted for life on land or at sea, and would have no hope for survival.
END_QUOTE
It has to be conceded that Gould was picking an easy target, one quickly demolished with a moment's thought. Polar bears spend a good amount of time in the water -- but don't seem, at least superficially, to be particularly optimized as water creatures. River otters are better off in this regard, and sea otters better still; sea lions take the level of adaptation to the water one step further, while true seals are so adapted to life in the sea that they find it difficult to get around on land. The whale is the endpoint of this spectrum, totally optimized for life in the sea, so much so that it will die if it goes onto land.
Of course, the variation from otter to seal to whale does not imply that otters and seals are ancestors of whales and dolphins -- nobody believes that -- but it takes little imagination to think that whales are descendants of partially-aquatic creatures along such broad lines, which modern fossil evidence has strongly confirmed.

Birds such as penguins have also returned to the sea. One of the fascinating observations of penguins swimming in a tank at a zoo is that they swim using their forelimbs, their wings, with their tail and feet playing strictly supporting roles. It takes little imagination to see the evolutionary principle of "leveraging off what's already available" in action: the penguin "flies" underwater in a fashion not broadly all that different from the way its ancestors flew through the air. Interestingly, although sea lions and true seals are closely related, they swim in different fashions, with sea lions (and walruses) obtaining propulsion from their front fins like the penguins, while seals use twisting motions of the body ending in the tailfins -- a neat example of the branching nature of evolution in action. Whatever works.
Similar thinking applies to the adaptation of animals to land. There are a number of different species of fish that can flop around on land and go cross-country, most notably the "walking catfish" of Southeast Asia that has been making a nuisance of itself in the American South since it was carelessly introduced there some years back.
The walking catfish doesn't do a particularly impressive job of getting around out of water. The froglike mudskipper, another Southeast Asian fish, is better adapted to land locomotion -- while amphibians like frogs are still better adapted to life on land, though as a rule they still must lay eggs in water.

There are cases where it actually is hard to see how there could have been a gradual progression of forms. Birds, bats, and the extinct pterosaurs have wings that are obviously modified legs, with the wings all derived from a pre-existing structure, as MET says they should be. It might have been literally "handy" for bats to have retained their front legs and grown wings on their back to look like tiny devils out of Dante, but evolution doesn't work that way. However, insects have wings that are clearly unrelated to legs. The general belief is that insect wings began life as a structure for another purpose, possibly a panel to absorb sunlight or radiate heat, with the structure proving useful for gliding purposes. Models have been constructed to show how it could happen, but they remain argued.
Such "indirect pathways" are called "pre-adaptations" -- though, since this term tends to misleadingly imply that some biostructure was deliberately obtained just because it might come in handy later, some prefer to use the more obscure but less loaded terms "exaptations" or "co-options" instead. One of the famous examples, promoted by Gould, is the "panda's thumb". A panda is a bear adapted to eating bamboo shoots -- an unusual practice for a bear, though given the prevalence of bamboo in Asia a practical habit -- and unlike other bears it has a "thumb" on its forepaws to help strip down the shoots for consumption. The "thumb" turns out to be nothing more than an extension of a normal bone in a bear's paw, the "radial sesamoid", that proved useful. The panda's thumb isn't much of a thumb, but it works.

Bears tend to be "handy" creatures, sitting or standing and using their forepaws as manipulators, and the panda has simply taken this process to a specialized extreme. In fact, in a sense the panda itself is something of an example of indirect pathways: who would have imagined a bear, a member of a family of carnivores, that eats grass for a living?
BACK_TO_TOP
