
* The early prokaryotic organisms that colonized the Earth acquired the ability to cooperate with each other in some cases. The ultimate result of a few of these "collaborations" was a new class of hybrid organism, the "eukaryotes", and a massive diversification of forms of life. The eukaryotes also introduced sexual reproduction, an adaptation that helped put evolution into a higher gear.
* The Archean gave way to the third of the four eons of the Earth, the "Proterozoic", about 2.5 billion years ago. Its introduction was marked by the first appearance of traces of oxygen in the atmosphere, obviously produced by cyanobacteria. By 2.2 billion years ago, oxygen was established in the atmosphere. This "Great Oxygenation Event (GOE)" wasn't that great, since the atmospheric concentration of oxygen was maybe a tenth, maybe less, than the current atmospheric concentration of oxygen of about 21%, by volume. That was still enough to kill off most of the anaerobic prokaryotes, since they were poisoned by oxygen. This is sometimes called the "first mass extinction event" – though it only left indirect traces in the fossil record.
The markers in the geological record of the GOE are distinctive: deposits of minerals older than 2.5 billion years that had been exposed to the atmosphere show no sign of oxidation, while oxidation becomes increasingly evident from that time on -- iron oxides, manganese oxides, hundreds of different oxides. The result was a slow mineralogical transformation of the surface of the Earth, with oxygen-laden ground water ensuring the transformation was more than merely skin-deep. The evolution would continue for a billion years, with a generation of minerals emerging, to provide feedstocks for a subsequent generation of minerals emerging from that.
At the time, there was no life on land, except possibly "pond scum" on the oceanic edges or blown inland by the winds. Given the prevalence of iron, the emerging continents acquired a red hue, from iron oxide. Oxygen helped boost weathering processes that slowly ground down rocky outcrops, resulting in growing sedimentary deposits.
The oxygen also began to form an "ozone layer" as solar radiation broke down the oxygen, O2, molecules in the upper atmosphere, forming ozone, O3. To that time, solar ultraviolet radiation had flooded down to the surface of the Earth, making existence difficult for such organisms as came above the protective surface of the sea. The ozone layer helped block the UV, though there was too little oxygen to more than lightly impede it.
For over a billion years, the Earth might well have seemed unchanging to any alien visitors that stopped by every now and then. The continental cratons shifted; new minerals arose; while single-celled organisms continued to evolve; but from space, it was unlikely that the planet's appearance changed much. The fossil record doesn't tell much about single-celled organisms, and so the Proterozoic isn't of so much interest to biologists -- indeed, biologists tend to simply lump the three first eons of the Earth into one block, the "Precambrian", as if to shrug it off.
Change started coming more rapidly late in the Proterozoic, about 850 million years ago. At that time, the movements of the continents had accumulated most of the cratons in a supercontinent named "Rodinia", surrounded by a global ocean named "Mirovia". Rodinia, however, then began to break up, a rift forming to shed off the Congo and Kalahari cratons -- which now form southern Africa. The West Africa craton then split off 800 million years ago.
By 750 million years ago, the breakup of Rodinia was in full steam, the supercontinent having split into north-south halves. The multiplication of coastline meant a multiplication of coastal erosion, with a comparable multiplication of oceanic mineral content and sediments. With more mineral nutrients available, photosynthetic microorganisms bloomed, increasing the oxygen content of the atmosphere. When these microorganisms died, they fell to the seafloor, to be buried in sediments -- sequestering their carbon, with the effect of a reduction in atmospheric CO2. The signature of this process is thick limestone deposits from that era, generated with the help of the dead microorganisms, with the deposits featuring a distinctive isotopic signature.
The algal blooms had the result of depleting atmospheric CO2. In addition, the breakup of Rodinia led to shallow inland seas. Water evaporating from these shallow seas contributed to more rainfall, and more erosion to cause the weathering of rock. The weathering of rock also absorbs CO2. However it happened, atmospheric CO2 concentrations fell. CO2 traps infrared radiation, and so a reduction in CO2 concentrations means less heat trapped by the Earth's atmosphere.
In an era when the Sun was dimmer than it is today, that had dramatic consequences, in the form of a global ice age. Glaciers leave clear marks on the terrain, with these marks extensively revealed in geological deposits dating from 740 million to 580 million years ago. They reveal that the planet largely or completely froze over during that time, forming a "Snowball Earth".
The ice shut down the bulk of photosynthesis and much of the CO2 absorption through weathering. Volcanic eruptions belched CO2 into the atmosphere, much as they always do, but the sinks to remove CO2 were shut down, with the result that CO2 concentrations climbed to higher and higher levels. The CO2 trapped solar heating, with the ice melting; as more dark land was exposed, it helped absorb more solar energy, boosting the heating.
The heating was boosted still further by the action of methane-producing microorganisms, methanogens, and possibly by methane produced from the Earth's mantle. Methane is an order of magnitude more potent a greenhouse gas than CO2, and large quantities of methane released into the atmosphere would produce dramatic warming.
Incidentally, the production of methane in the mantle can also lead to production of more elaborate hydrocarbons. There have been suggestions that the Earth's oil was mostly produced from the mantle as well, and not from ancient deposits of biomass buried in the Earth. The notion of "abiotic oil" is not the conventional wisdom; it is not seen as an unreasonable idea in itself, but cranks are inclined to treat it as a solidly-established fact. The usual rationale for doing so is to suggest that oil supplies are effectively unlimited.
In any case, once the snow had melted off the ground, CO2 absorption by weathering started up again, as did algal blooms. Methane breaks down spontaneously into CO2 and water on a half-life of a few decades, so it didn't persist for very long after production fell off. The end result was the Earth rolled downhill towards cooling again, with the cycle repeated a number of times. Exactly how many is a matter of argument, and in fact it's a matter of argument that the Earth ever froze over completely. Climate modelers have found it difficult to come up with simulations in which the planet completely freezes over, with a faction countering "Snowball Earth" with "Slushball Earth". The discussion continues.

The cycles of algal blooms did have one persistent effect of building up oxygen concentrations in the atmosphere. When the Earth got hot, the algae pumped out oxygen; when the planet froze down, the oxygen concentration remained roughly constant. By about 650 million years ago, oxygen concentrations had risen to near-modern levels, in a second Great Oxygenation Event, with a protective ozone layer providing a shield against most solar ultraviolet.
BACK_TO_TOP* The "eukaryotes" seem to have emerged from the prokaryotes about two billion years ago, in the wake of the first GOE. The eukaryotes are about ten times bigger in diameter than prokaryotes, and have a central "nucleus" that contains their genetic material. They also have an elaborate set of cellular "organelles". The "mitochondrion" of most eukaryotic cells is used in energy processing, and has its own little distinct genetic code, separate from the main cellular code in the eukaryotic nucleus. Plants add an organelle called a "plastid" or "chloroplast", which performs photosynthesis, and has its own little genome. The genomes of these organelles are no longer sufficient to direct all the organelle's functions, with the genome of the organelles interacting with the genome of the host cell to get the job done.
It is now accepted among biologists that the eukaryotic cell arose as a mutualistic or symbiotic relationship between various prokaryotes. The basic idea was originally proposed by a Russian biologist named Konstantin Merezhovsky (1855:1921) and fleshed out by an American biologist named Ivan Wallin (1883:1969) in the 1920s. It was generally regarded as a wild idea, and it certainly wasn't well supported by the evidence available. The modern understanding of DNA and molecular genetics opened the door to a more detailed examination of the matter, and in 1963 an American biologist named Lynn Margulis (1938:2011) made a case for the idea that eventually proved persuasive.
In fact, although some have claimed the symbiotic origin of the eukaryotic cell was a wild improbability, the evidence suggests that it was almost inevitable. As Margulis pointed out such symbiotic associations clearly took place more than once, since plant cells have both mitochondria and chloroplasts, and there may have been other events as well: there are even some strains of bacteria that appear to be symbiotic hybrids of earlier strains. Symbiosis often starts from parasitism, the parasite then acquiring a more benign relationship with its host.
The eukaryotic host cell seems to have been derived from the archaea -- in fact, from a specific lineage of the archaea, with some taxonomists pointing out that makes the eukaryotes a subclade of the archaea. As for the mitochondrion, it appears to have come from the proteobacteria, while the chloroplast from the cyanobacteria. Symbiotic relationships between different prokaryotes are actually perfectly ordinary in the modern world, and some of these relationships are much along the lines of those believed to have given rise to the eukaryotes.
The eukaryotic nucleus may have initially functioned as a barrier to reduce transfers of genes from the symbionts to the host cell genome, which can play hell with the host genome. Genetic analysis does show that there seems to have been some gene transfer between mitochondria and the eukaryotic host genome; in fact it is becoming increasingly apparent that gene transfers occur surprisingly often in eukaryotes, though not remotely at the level at which they do in prokaryotes.
The eukaryotes would ultimately branch out into true multicellular organisms. As with horizontal gene transfer among prokaryotes, the symbiotic origins of the eukaryotic cell tend to complicate the branching tree of life. Some creationists have claimed that this complex of roots completely undermines the notion of evolutionary common descent, but though it does add fine print, common descent remains a completely valid concept above the level of microorganisms. The fine print only amounts to an expansion of the evolutionary vision, not a refutation of it.
Margulis, along with her son (with astronomer popstar Carl Sagan) Dorion Sagan (born 1959), pushed the idea that symbiosis is actually the primary source of variation for evolution, claiming that her ideas fundamentally changed the rules of evolutionary science. In this, Margulis was not taken very seriously -- partly because she was a noisy and abrasive contrarian, not merely sniping at her colleagues in the evolutionary biology community, much to the glee of creationists, but also obsessively embracing far-fetched conspiracy stories and any other challenge to sensibility that caught her attention.
More significantly, while symbiosis is clearly a very important evolutionary process and Margulis deserved great credit for highlighting it, as Ernst Mayr pointed out there is little evidence in the species diversity of modern macro-organisms to suggest that this diversity arose directly from symbiotic processes. Margulis was particularly mocked for promoting a paper by Donald Williamson, a retired British zoologist, who claimed that caterpillars and butterflies were originally separate lineages, but became one line through hybridization. The most tolerant response was that it was an interesting idea to examine -- but in any case, it didn't stand up under examination.
* The eukaryotes were an extremely significant development in the history of life. Energetic processes in prokaryotes are dependent on the cell membrane; as the prokaryote gets larger, the volume increases well more rapidly than the surface area, and so as the prokaryote grows, it finds it ever harder to stay fed. Eukaryotic cells, in contrast, use mitochondria to support energetic processes, and so they can grow much larger, giving them far more options.
The rest of this survey concerns eukaryotes, but that shouldn't be interpreted as dismissing prokaryotes as unimportant in any sense. There is a vast range of diversity among the prokaryotes, and we're still learning much about them -- as noted previously, we've tended to focus on pathogens while ignoring the far larger number of benign microorganisms. Prokaryotes have been around for a long time, and calling them "simple" can be misleading. All organisms are highly elaborate, and in many cases the prokaryotes are capable of performing some very intriguing tricks. Much of the Earth's biomass is prokaryotic, and in fact there are about as many prokaryotic cells in our bodies -- mostly in the form of intestinal flora -- as eukaryotic cells. Even if the prokaryotic cells are about a thousand times less massive, that's still a revelation.
BACK_TO_TOP* The eukaryotes can be divided into single-celled organisms, generally referred to as "protists", and multicellular organisms, or "mega-eukaryotes". The mega-eukaryotes include the fungi, plants, and animals. There are also some other mega-eukaryotes that are sometimes categorized as protists, which hints at the difficulties in determining the ancestry and relationships among the eukaryotes -- some single-celled eukaryotes being more closely related to some mega-eukaryotes than they are to other single-celled eukaryotes.
The term "protist" only defines a grade, not a clade; taxonomists actually break the protists down into about 20 different kingdoms, the number being a subject of argument. The protists include some members that are photosynthetic, while others are heterotrophs -- though in keeping with the nature of the group as a grade, some heterotrophic protists are closer to some photosynthetic protists than they are to other heterotrophic eukaryotes. In fact, there are groupings of eukaryotes that include both.
The earliest of the modern protists date back to about 1.2 billion years ago, and are now bewilderingly diverse. While some protists reproduce by fission, sexual reproduction is common among them as well. Some that live in anaerobic environments, such as protists included in our intestinal fauna, don't have mitochondria, performing metabolism by methods unique to themselves. Some can form spores. Some have flagella to get around; some are ringed with hairs or "cilia" for mobility; and some, the "amoebas", are flowing blobs.
* The most prominent example of photosynthetic protists are the "golden algae". They are generally found in bodies of fresh water, though some are found in the oceans, with oceanic variants featuring silicate skeletons. Most of the oceanic "diatoms" are also photosynthetic. They are distinguished by their intricate and elegant silicate "tests" or "shells".
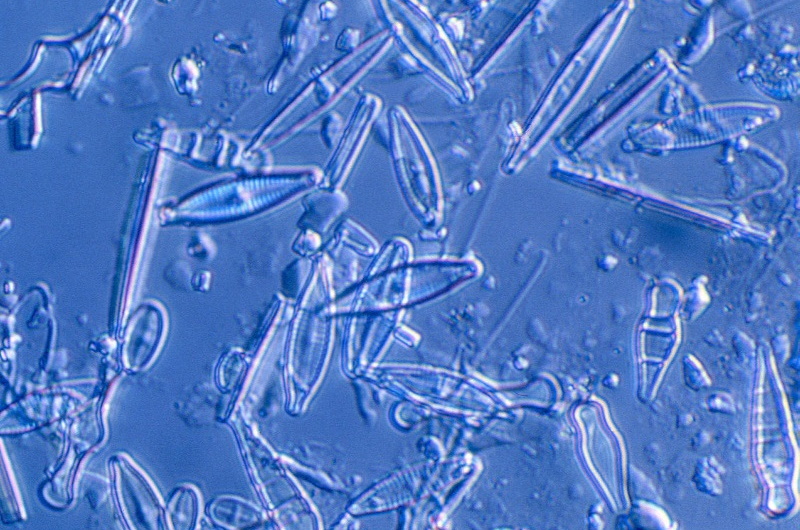
Protists like the "oomycotes" are heterotrophs that feed externally on decaying matter or act as parasites, throwing out threads known as "hyphae" that secrete enzymes to digest the external matter. This is the distinguishing characteristic of the fungi, and it was once thought the oomycotes were fungi. The two kingdoms are, however, very distinct biochemically, though oomycote species are often given names like "rusts" and "mildews" that would seem more appropriate for fungi -- in fact, the group is sometimes still referred to as the "water molds". The infamous oomycote Phytophthora was the "potato blight" that cause the ghastly Irish potato famine of the 1840s that depopulated the country through starvation and forced emigration.
The "cellular slime molds" are, as their name suggests, also funguslike. They are amoebic in form, though as discussed later they are capable of some very interesting collective behavior. The "true slime molds" AKA "plasmodial slime molds" are also funguslike, but they are very different from the cellular slime molds. The true slime molds are single-celled, but the cell can be extremely large by cellular standards, and it will have multiple nuclei.
There are a number of groups of animal-like protists -- heterotrophs that actually ingest food. The animal-like protists are known collectively as "protozoans", though once again, that's just a grade. They include:
It should be noted that anyone who claims the fossil record is sparse hasn't considered the radiolaria and the forams: they form layers under the seabed, and some tropical beaches are made up of almost nothing but their shiny white remains. Their evolutionary history is laid out almost like the pages of a book, the only problem being that such organisms are hard to study in the lab, and our knowledge of the lifestyles of living species is incomplete. The radiolaria and the forams are one of the "markers" used to identify distinct geological layers, and indeed forams are used by oil companies to identify strata that could be associated with oil deposits.

It should also be noted that fossil evidence for protists only goes back about 800 million years, and that not all the modern groups, for example foraminifera, necessarily go back that far. Just like more elaborate organisms, they have evolved in various directions, with new forms arising and old forms dying out. Popular science literature sometimes refers to modern organisms that can be presumed to be similar to ancient ones as "living fossils", but that's a bit of a loaded term. All organisms living today are descendants of ancient ancestors, and if some are very much different from their ancestors, that doesn't imply that others that haven't changed as much are simply carbon copies of their forebears. No examples of living species are parents of another species: they are all cousins, though some certainly may look far more like the grandparents than others.
BACK_TO_TOP* While the bulk of protists reproduce asexually by division, some do reproduce sexually, with two parents providing half their genome to create a new generation with a hybrid genome. It is generally believed that the sexual reproduction emerged with the protists, and it's worthwhile to take a general closeup of the subject of sex because of its importance in evolutionary science.
There has been a long debate among evolutionary biologists over the relative merits of asexual and sexual reproduction, but few dispute that by the evidence, the rise of sexual reproduction was a major event in the evolution of life. In fact, it is a troublesome oversimplification to say that mutational change drives evolutionary processes, since the "recombination" of genomes through sex is a major factor in promoting genetic variability, in essence rearranging the "teams" of genes with every generation, with the effect of accelerating evolutionary change. If a particular mutation gives an advantage to one member of a species and another mutation gives an advantage to a second member of that species, then progeny of those two organisms can in the best case get a doubled advantage. In a sense, the evolution of sex was an example of the "evolution of evolution".
However, sexual reproduction is more than a bit puzzling. Evolution is not an end in itself; as noted in the discussion of punctuated equilibrium, a well-adapted organism may well continue to propagate for a long time, with only relatively minor or obscure modifications that do not change its basic form and habits, until new selection pressures begin to affect its fat and happy existence. Evolution occurs in response to selection pressures -- it's much more driven than a driver. In addition, since sexual reproduction involves a fair amount of overhead, the selective advantage provided by sex had to be significant, or it wouldn't have caught on. Asexual production permits rapid and efficient replication, and if the survival of species was driven strictly by the ability to reproduce, asexual reproduction would win hands-down. From the point of view of the "selfish gene", asexual reproduction ensures the maximum propagation of the "winning team" of genes.
There are a number of different, and not necessarily mutually exclusive, ideas as to why sexual reproduction persists. One long-standing notion is that it allows organisms to "clean out" a gradual buildup of deleterious mutations by reshuffling the genomes provided by the two parents and selecting the winners. A more popular notion, devised by William Hamilton, is called the "parasitic Red Queen's race" -- after the animate chesspiece in Lewis Carroll's THROUGH THE LOOKING-GLASS who had to run as fast as she could just to stay in the same place.
The idea is that organisms featuring sexual reproduction tend to be relatively long-lived, with much longer generation times than the asexual pathogens that infect them. The fast-breeding pathogens are likely to mutate and acquire some means of breaking through the defenses of their hosts in time, in a sense "breaking the combination on the lock" of those defenses. Sexual reproduction ensured that the hosts could scramble the "combination" and stay just one step ahead of the pathogens; importantly, the genes in control of the mammalian immune system are highly polymorphic, ensuring that there is a wide range of "combinations". Even when a pathogen manages to crack the "combination", it will not be able to take down the entire population of a genetically-diverse species.
In other words, the fitness of large organisms is continually being eroded by pathogens, the "winning team" will not go on winning indefinitely, and the large organisms have to keep up. From the point of view of the genetic imperative, asexual reproduction would be better, but the problem is that pathogens will move in on that racket sooner rather than later, and so organisms that survive rearrange their teams of genes on a regular basis instead. The science-fiction notion of a "satan bug" that could take out every single human on the planet is, or at least we have good and hopeful reasons to judge it is, a fantasy.
An interesting example in this case is that of the modern cultivated banana. Wild bananas have fruit with large seeds, while cultivated bananas are mutants with vestigial seeds. Cultivated bananas can only be crossbred in the lab, and it requires considerable effort. Commercial production of bananas involves the replanting of cuttings -- or in effect the cultivated banana reproduces asexually, with humans as the agent of reproduction. There are a number of different varieties of cultivated bananas, such as the starchy plantain, but outside of the tropics the selection of bananas available to consumers is very narrow. Up to the 1950s, the primary commercial banana was the "Gros Michel" AKA "Big Mike", which was all but wiped out worldwide by a fungal infection. The current commercial banana is the "Cavendish", and it is now being threatened by a mutant of the fungus that wiped out the Gros Michel. The asexual cultivated banana is at an inherent disadvantage in the Red Queen's race with its fungal parasite.

* Of course, asexual production does persist. Dandelions generally reproduce asexually -- the flower is vestigial -- and get away with it by having short, rapid life-cycles, breeding wildly before pathogens infect them and kill them off. Some organisms have phased sexuality, reproducing asexually for a few generations and then, under some circumstances, reproducing sexually. Other organisms that are normally sexual reproducers may have a species among their ranks that reproduces asexually -- one interesting example being an asexual species of crayfish that has been adopted by commercial fish farmers -- though such species tend to be unusual since they are vulnerable to being culled out by pathogens.
Lab experiments with microorganisms that can either produce sexually or asexually show they will reproduce sexually if their living conditions often change; once conditions are left constant, asexuality tends to take over again. However, outside of the lab, species are confronted with pathogens that are continuously evolving into new threats, maintaining continuous pressure against asexual reproducers -- ensuring that they are forever doomed to remain a minority, at least among large organisms with long life-cycles.
Trying to figure out the variations on the theme of asexual versus sexual reproduction has been a challenge to biologists, and in fact sexual reproduction itself involves a number of intriguing puzzles. There is no doubt it emerged in phases: bacteria, as mentioned, have a limited ability to transfer genetic information, and it seems plausible that a scheme along this line could have led ultimately to sexual reproduction. Some speculate that the diploid cell, the basis for sexual reproduction as we generally understand it, may have actually well preceded the origins of sex, being the product of some error of replication of a haploid cell. With dual sets of chromosomes, the new diploid cell was prone to disastrous errors in replication, until meiosis emerged and finally set things right. Sexual reproduction emerged as the next step.
There is also the puzzle of how male and female organisms could have emerged in lockstep, but that puzzle turns out to be easier to toss out. At the outset, such organisms would likely have been hermaphroditic -- that is, each both male and female. That's nothing unusual among larger modern organisms, snails being a well-known example. That still leaves the question of how two distinct sexes emerged, but one could imagine hermaphrodites that cycled between genders, ultimately ending up with the gender fixed at birth.

That leads to another puzzle: why two distinct sexes? Why aren't hermaphrodites the norm? It would seem that a species would reproduce more effectively if all its members were capable of bearing young. Hermaphrodites can also inseminate themselves, though "selfing" tends to lead to unhealthy in-breeding and hermaphroditic species tend to have mechanisms to prevent it when they can find partners. However, from an evolutionary point of view selfing is much healthier than dying out.
The question of males and females turns out to be particularly devious. Traditionally, the explanation was that it made sense to have males who generate their sperm rapidly and scatter their genomes readily, as well as females who actually have to deal with the overhead of bearing young. The first problem with this notion is that, while it is "cheaper" to generate one sperm than one egg, it usually takes far more than one sperm to inseminate one egg. In fact, since the female reproductive system can be hostile to sperm, it may take millions of sperm to inseminate one egg.
Such hostility is puzzling, since it is in the evolutionary interests of species to encourage their own reproduction. One possible answer is that the female reproductive system is necessarily very "nurturing", which means that it's also a nurturing environment for pathogens. That results in an evolutionary race to raise the barrier of entry to pathogens without denying access to sperm, and so the barrier gets raised to whatever level that the males can still overcome on a normal basis. In any case, it isn't necessarily easier, at least from a broad point of view, to make sperm than it is to make eggs.
In addition, in many species the males do most of the work of child-rearing, one of the extreme cases being the seahorses and their elongated relatives, the pipefish, in which the female lays her eggs in a pouch provided by the male -- the female has a "penis" or ovipositor to do the job -- and the babies come to term in the male.

The classic argument for the existence of males and females is not entirely wrong. Imagine a horror-movie scenario in with a pathogen was introduced that decimated the human population, and suppose it was selective with regards to sex. If it killed off most of the women, then from the point of view of rebuilding the population, it might as well have killed off most of the men too -- since the surviving women would effectively breed no faster than they would have if as many of the men had been killed off. In contrast, if it killed off most of the men, the population would be rebuilt within a generation. Could one healthy man keep ten fertile women pregnant? Sure. A hundred women? Possibly, though it might be somewhat exhausting. The surviving males would win the genetic jackpot.
However, the case would be exactly the opposite for seahorses. So then, why males and females? There may actually be a simpler explanation, and it relates to the fact that bisexual reproduction hasn't displaced hermaphrodism.
The traditional concept is that hermaphrodism occurs in immobile species -- plants or sessile animals -- or species that aren't highly mobile, such as snails. In such circumstances, coming into sexual contact with another member of the same species is relatively unlikely, and having two different sexes would halve the odds of doing so. Being hermaphroditic guarantees that any adult member of the same species is a candidate sex partner. Flowering plants are generally hermaphroditic, since they are sexually dependent on symbiotic pollinators like bees, hummingbirds, and the like; it might not be impossible for a pollinator to acquire different behaviors to handle two distinctly different plant genders, but it would certainly make things more difficult.
This scenario clearly has some truth in it, but it's also not the complete truth, because there are plenty of hermaphroditic species that can get around perfectly well. Possibly the answer to the puzzle of bisexual reproduction versus hermaphrodism is that they are just two systems that happen to work. As noted, bisexual reproduction likely originated from hermaphrodites, and in fact there are existing species where the two "genders" are hermaphrodites and males. The hermaphrodites can reproduce among themselves, or can reproduce with males.
The males, from this point of view, are "broken" hermaphrodites; further "breakage" could produce females. If the hermaphrodites were at a small selective disadvantage from having to maintain two reproductive systems, they would gradually be eliminated from the gene pool. It's much more straightforward for bisexual reproduction to evolve from hermaphrodism through the loss of the reproductive system by halves, than for bisexual reproducers to evolve into hermaphrodites by developing a new reproductive system. Bisexual reproduction is likely to emerge from hermaphrodism on occasion just by the odds.
It is not so hard to figure why there are usually roughly equal numbers of males and females in a given bisexual species. It's a question of market saturation: if more human males were born than females, given that humans are basically monogamous, the excess males would simply "go to waste", and from an evolutionary point of view they would not be a good investment of effort. Humans are of course only one species, but theoretical analysis has showed that the optimum ratio of males and females for any bisexual species is, under stable circumstances, 50:50.
It might seem that harem animals, such as antelopes or seals, in which only a fairly small portion of the males ever get to reproduce, imply a violation of the 50:50 rule. In fact, mother seals bear roughly as many males as females. Even though most of those males won't reproduce, the parents still have an incentive to commit resources to the game in the hope of winning. A harem system is simply a modification of the sex contest in which fewer prizes are handed out, but the prizes are bigger. It is interesting, however, that in harem-oriented species the males tend to be bigger as adults than the females, with the difference in size growing with the average size of the harem: bull elephant seals have unusually big harems, and are monsters compared to the cows. The emphasis on size is not surprising, considering that under a harem scheme it is literally only the biggest bullies that get to sire offspring.

A fourth, somewhat absurd, question about sex is: why not have three or more sexes? The notion gets played up in sci-fi stories on occasion, usually as a gag -- on a planet with a dozen sexes, scoring on a Saturday night would be hideously troublesome. In reality there are organisms that have been described as having more than two sexes, in fact sometimes hundreds of them, though some purists prefer to use the term "mating types" instead of "sexes" because they're not "sexes" as most would think of the term: all members of such species have effectively the same reproductive gear.
Such "isogamous" organisms in essence produce eggs, but not sperm; true slime molds are a prominent example. The only way in which such organisms have sexes is that the gametes of different individuals, though structurally identical, are biochemically "keyed", partly to avoid self-insemination. This creates an evolutionary pressure to keep adding sexes: if there were gametes of types A and B that could breed with each other, then there would be an advantage to evolving a type C that could breed with A and B. By the same logic, there would be an advantage to a type D that could breed with A, B, and C ... and then to type E, and beyond.
Even consideration of this fine print still reveals that no organism has more than two genetic parents, no matter how confusing the sexual arrangements of the parents are in practice. The reason is that sexual fusion of two germ cells into a composite zygote cell is complicated. This is why there's the female egg / male sperm asymmetry in sexual reproduction: it's simpler to combine the complicated egg with minimalist sperm and get a working hybrid than it would be to hybridize two germ cells as complicated as eggs, with all their cellular organelles. Isogamous organisms are not an exception to this constraint: the rules of sex between the different types of gametes determine which one of the partners will provide the cellular organelles.
Trying to get two germ cells to mesh together is tricky enough. Trying to hybridize three germ cells of any configuration and get something that works would be nightmarishly complicated. An evolutionary path to such a complicated scheme would be much more elaborate than the path that led to two sexes, and it's hard to identify any advantage of three sexes over two that could provide selection pressures to drive evolution along such a path. Three or more sexes really are troublesome.
* Incidentally, the 50:50 gender rule has some fine print -- it actually means that the optimum level of relative investment in males versus females is 50:50. If males require as much investment as females, then the number of males matches the number of females; but in a species where males are twice as easy for a mother to produce as females, then the optimum ratio of males to females is predictably 2:1: "Two for the price of one!".
There are also some creatures that produce a few males and a large number of females -- but the brothers and sisters breed among themselves and such creatures can be seen as "virtual" asexual reproducers, clearly being descendants of sexual reproducers that "rediscovered" some of the benefits of asexual reproduction. In fact, in some species of mites, the offspring hatch inside the mother mite's body, with the male inseminating his sisters before they eat their way out of their unfortunate mother, the male then promptly dying. The female mites are "born pregnant", and on a casual inspection such a species might actually appear to be an asexual reproducer -- though in reality, it has taken a roundabout evolutionary route to get back to a bastardized approximation of where its distant ancestors started from.
Another strange tale of sexual reproduction is that of the fish known as the blue-headed wrasse. Only the males are blue-headed, the females are dull-colored. They are a harem species, with one male controlling a school of females. However, as mentioned previously, some fish can change sex during their lives, and the blue-headed wrasse is one of them. If the male in control of the harem dies, the biggest female becomes a male and takes over. It's the best of both worlds.
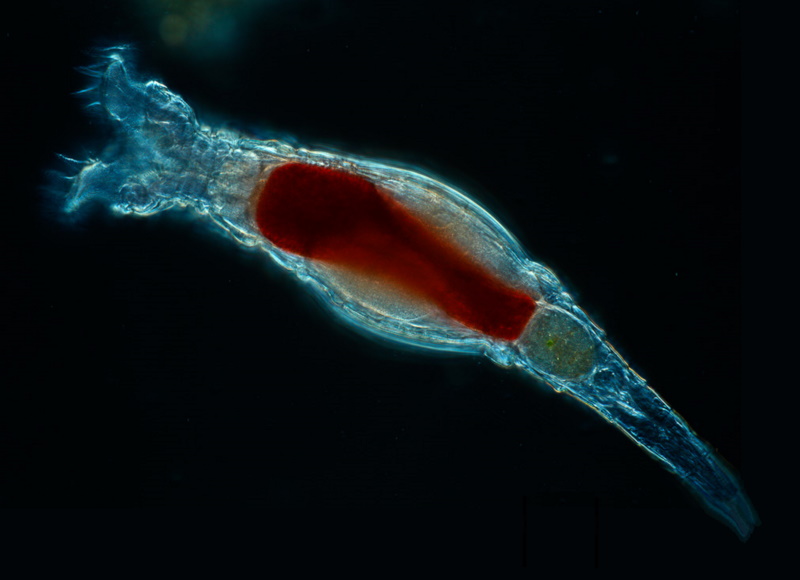
Such tales hint at how complicated the matter of sex really is, and it must be emphasized that the discussion here is brief and glib -- not everyone agrees with the "parasitic Red Queen's race" idea, for example. There are special cases where it just doesn't seem to work, the most prominent case being the protists known as "bdelloid rotifers", all of which have been asexual reproducers for at least 85 million years, and which seem to be thriving.
Investigation of the bdelloid rotifer genome hints that they get by on horizontal gene transfers, with 10% of their genome lifted from bacteria, fungi, and plants. How they perform this trick remains a puzzle; it appears to have something to do with the fact that, without water, bdelloid rotifers dry out, with their dessicated form reactivated when water returns. The reactivation, it seems, encourages pickup of foreign genes. Sex turns out to be one of the most devious topics in evolutionary science, and it's likely to provide job security for biologists for a long time to come.
BACK_TO_TOP
