
* The confusion into which evolutionary theory fell in the late 19th century did not last. In the early 20th century, the discovery of radioactive dating and the rediscovery of Gregor Mendel's work would do much to dispel the confusion.
* Creationists have always proclaimed the imminent death of evolutionary science, though the obituary has so far proven somewhat exaggerated. However, in the year 1900, it was as close to dead as it has ever been. Although the idea of evolution was now scientific dogma, arguments prevailed, with few naturalists agreeing about what really drove evolution.
The first key to the revival of evolutionary theory was the discovery of radioactivity by the French physicist Anton Henri Becquerel (1852:1908), who found that some materials emitted a mysterious radiation that would fog photographic film. In a few years it would be understood that radioactivity was caused by the energetic breakdown of the atoms making up the materials, and that the energies produced by such breakdowns were huge -- as given by the famous formulation of the German-American physicist Albert Einstein (1879:1955), "E=MC^2".
The physicists had discovered an energy process that allowed the Universe to be much older than Lord Kelvin had asserted. In addition, since different radioactive materials have different and highly specific decay rates or "half-lives", radioactivity gave geologists a tool to actually estimate the age of many mineral samples. Not only was the Universe now old enough to allow evolution to take place, it was also possible to provide reasonably hard dates on the fossil record.
* The second key rested in the first steps towards the understanding of heredity. It had been known long before Darwin's time that organisms were made up of discrete "cells", and that specific "germ cells" -- eggs and sperm -- were the "seeds" of generation of an organism. Organisms such as plants and animals have cells with distinct subordinate "organelles" and a walled-off central "nucleus". By the 1880s, microscopic studies seemed to strongly hint that the mechanisms of heredity were associated with the cell nucleus. Unfertilized eggs and sperm carried "pronuclei", and observations showed that a sperm was little more than a pronucleus with a tail. A fertilized egg contained two pronuclei that quickly merged, to be followed by cell division and construction of the organism.
In 1882, a German biologist named Walther Flemming (1843:1905) published a report concerning his observations of a set of "threads", later called "chromosomes", in salamander larval cells, that seemed to divide along with the cell. New staining techniques and further improvements in microscopy brought the chromosomes into closer view, and showed that the chromosomes were duplicated before cell division. The chromosomes were suspected to be structures supporting the machinery of heredity, but nobody knew just how. Some believed that each chromosome contained a complete blueprint for the organism.
There is no evidence that Gregor Mendel heard of this discovery before his death two years later. Progress forward depended on a Dutch botanist named Hugo de Vries (1848:1935). The Dutch are big on raising flowers and in the mid-1880s, de Vries began to investigate the breeding of flowers, leveraging off the statistical methods developed by Galton -- they were perfectly valid in themselves, and a particular example of Galton's wildly mixed legacy. By that time, it was known that the cellular chromosomes were involved in cell replication: they existed in pairs that split up in the germ cells of parents, a process known as "meiosis", and then recombined in the fertilized egg cells of progeny.
De Vries and others believed, at the time on somewhat circumstantial evidence, that the chromosomes were carriers of hereditary information. Nobody had any clear idea of the details, but de Vries suggested, again correctly, that different traits were found as "units" positioned on the chromosomes. He had no idea of the precise nature of these "units", but he did give them the name "pangenes", in homage to Darwin's theory of pangenesis. De Vries, a focused and humorless man, pursued the concept through statistical analysis and plant cross-breeding experiments, and gradually came upon the idea of traits being passed from each parent in the form of pangenes -- resulting in the 3:1 ratio of traits in descendants as observed by Mendel.
De Vries published this conclusion in 1900. He was aware of Mendel's 1866 paper, but didn't cite Mendel as a source. A German botanist named Carl Correns (1864:1933) had also been performing plant crossbreeding experiments and had come to conclusions similar to those of de Vries -- but Correns was quick to point to Mendel's priority.
It was difficult to say that the issue was highly controversial at the time, the revelation not being seen as revolutionary for the moment -- though in fact, it was. However, the end result was that Mendel's work had been fished out of the pigeonhole in which it had been gathering dust for over three decades. Not everyone was impressed with Mendel's work, some even calling it fraud, but others were intrigued and worked to continue Mendel's research. A Danish botanist named Wilhelm Johannsen (1857:1927) gave the then-hypothetical unit of heredity the name "gene" (by stripping down "pangene"), which the British biologist William Bateson (1861:1926) used to mint the word "genetics" for the study of heredity.
This is not to say that the confusion had been resolved. Many of the early "geneticists" like William Bateson didn't believe that genes physically resided on chromosomes, instead adopting fuzzy ideas about energy states or waves or whatever, not coded arrangements of molecules. A German biologist named Theodor Boveri (1862:1915) and an American biologist named Walter Sutton (1877:1916) suggested otherwise, pointing out the strong connection between the "pairing" of genes with the division and then recombination of chromosomes, but for the moment the issue remained arguable.
BACK_TO_TOP* It seemed at the time that the confusion over evolutionary science was simply changing its terminology somewhat, but now the final arbiter of scientific disputes, cold hard data, intervened, in the form of careful studies performed by a meticulous American experimentalist, Thomas Hunt Morgan (1866:1945), at Columbia University in New York City.
Morgan was a pleasant gentleman from Kentucky, a nephew of Confederate cavalry raider John Hunt Morgan -- though Thomas never knew his famous uncle, General Morgan having been gunned down by Union troopers late in the Civil War. Thomas Hunt Morgan was a thorough "empiricist", focused on experimental data, with little respect for either religion or for the fuzziness of Darwinian thought as it existed at the time. Polite in his demeanor, he still mocked the vague hereditary ideas of the era as "invisible germs" whose sole functions were those "imagination bestows on them". Morgan wanted something more tangible, and he found it in the unlikely form of the little Drosophila melanogaster fruitfly.
He was cross-breeding fruitflies in his lab at Columbia by 1909. He found the flies "wonderful material -- they breed all year round and produce a new generation every twelve days." They were much better subjects than pea plants or flowers, permitting him to observe a hundred generations in less than four years. It was, however, untidy work, with the flies raised in rows of small milk bottles on rotten bananas in his "fly room" at Columbia, resulting in a rank-smelling environment buzzing with little escaped flies.
Morgan was initially skeptical about Mendel since it seemed as though traits were blended in progeny, and the business of Mendel's 3:1 ratio was contradicted by the fact that in most animal species, males and females were born in roughly equal numbers -- gender didn't appear to be based on paired dominant and recessive traits, so why should anything else be? Morgan was just looking for mutations, and did not expect to validate Mendel.
After about a year of effort, the lab crew produced their first distinctive mutant -- a male fly with white eyes, instead of the normal red eyes. He crossbred the mutant male with a red-eyed female, then crossbred the progeny. All of the first generation had red eyes -- but the second generation featured both red eyes and white eyes, in a ratio of 3:1. In addition, all the white-eyed flies were male, without exception, and so it appeared the white eye color trait was linked to the hereditary mechanism that determined Sex. Two more mutants were soon discovered: yellow body color and miniature wings. Both bred in a 3:1 ratio, and both were sex-linked. Over the next few years, dozens of mutations were uncovered; Morgan and his students were observing evolutionary variation in action.

The various mutations fell into four groups, with each group apparently being correlated with a particular chromosome. The only problem was that there were rare exceptions, such as very unusual cases of white-eyed female fruitflies. Such an inconsistency promised to make life difficult for the chromosome theory of inheritance, but Morgan discovered an out. In 1909 a Belgian cytologist (cell researcher) named Frans Janssens had observed that during meiosis, chromosome pairs would perform a "crossover" at certain locations, to break apart and then reform.
Morgan deduced that the genes that he presumed existed were strung like beads on a string along a chromosome, and that the recombinations of chromosomes were likely to switch genes around. Furthermore, the likelihood of two genes on the same chromosome being separated from each other was obviously greater the farther they were apart -- that is, if they were close together then they were likely to be exchanged together, but if they were far apart one would be exchanged and the other left behind. This discovery turned out to be much more than a mere loophole: creation of "linkage maps" allowed Morgan and his students to actually locate genes on the chromosomes. However, at the time, they couldn't provide the slightest detail of what a gene actually was.
Morgan had validated Mendel without intending to, and became a believer in Mendelian genetics, as well as the chromosome theory of inheritance. He published his results in 1915 in THE MECHANISM OF MENDELIAN INHERITANCE, co-authored with three of his students, Alfred Sturtevant (1891:1970), Calvin Bridges (1889:1938), and Hermann Muller (1890:1967). Morgan won the Nobel Prize for his work in 1933. His three students would go on to become prominent geneticists in their own right, with Muller winning the Nobel Prize in 1946.
* Incidentally, the early detailing of mutational processes by Thomas Hunt Morgan and his people led to a scientific dispute that went off the rails. Morgan believed that new mutations were a significant driving force in evolution; others disagreed, insisting that existing variation in a population was the feedstock of natural selection. Continued research would generally vindicate Morgan, and today the importance of new mutations in evolutionary change is the accepted wisdom.
The irony is that Morgan is still widely perceived as being on the blindly wrongheaded side of the argument, thanks to misrepresentations by his adversaries that claimed Morgan simply discarded selection completely, and asserted evolution resulted strictly from sequences of mutations. In other words, they portrayed Morgan as proposing nothing more than a variation on saltationism, with the lucky set of mutations occurring in sequence instead of all at once. In reality, Morgan never proposed any such thing -- but sneers at his foolish "mutationism" still linger in the literature.
BACK_TO_TOP* The acceptance of the Mendelian chromosomal theory of inheritance provided a wide range of insights, in particular in nailing down some of the puzzles of hereditary diseases. The classic example is the case of the "hemophilia" that was passed down to some of the male descendants of Queen Victoria. Hemophilia is a syndrome in which the blood-clotting mechanism of a victim is dysfunctional from birth: a simple scratch may cause him to bleed to death. The use of "him" is significant, since only the male descendants of the queen suffered from the syndrome.

Most cells of a human being have 23 pairs of chromosomes -- due to the pair organization, a normal human cell is said to be "diploid". In normal cell duplication, the 23 pairs of chromosomes are completely duplicated, this process being known as "mitosis". In the formation of germ cells -- sperm in males, eggs or ova in females -- the 23 pairs are divided into two half-sets, a process known as "meiosis" that creates a "haploid" cell. As Frans Janssens had observed, in most sexual species the process of meiosis doesn't simply grab one of the chromosomes in each pair -- it takes the pair and through crossover uses complementary parts of both to assemble a new equivalent chromosome, effectively scrambling their genetic contributions.
When a male's haploid sperm cell joins up with a female's haploid ova, the 23 half-pairs provided by each parent join up to provide 23 full chromosome pairs, resulting in a diploid "zygote" with half of its genetic complement provided by each parent.
22 of the human chromosomes are symmetrical, with coding differences but the same general structure between the pairs. These symmetrical chromosomes are called "autosomes". The exception is the pair consisting of the "X chromosome" and the "Y" chromosome, named because of their four-armed and three-armed arrangements respectively. A male human has an X and a Y chromosome; a female has two X chromosomes. It should be noted that not only do the X and Y chromosomes differ from the rest of the chromosome pairs in having different configurations, they also differ in only partly crossing over during meiosis.
The male determines the gender of the child, since the male's sperm will provide either an X or a Y chromosome, while the female's egg will only have the X chromosome. Although genes are of course involved in the appropriate construction of a male or female child, sex selection is actually at the chromosomal level -- which is why gender follows its own special, simple law of inheritance. Incidentally, in birds the scheme is exactly the opposite: ZW gives a female, ZZ a male, hinting at the fact that sex determination in general is a complicated matter, with highly variable rules. Not only are some species asexual or hermaphroditic, in some sexual species the gender is not fixed at conception, for example with the sex of many species of reptiles determined by the temperature of the egg. In certain species of fish and other organisms, an individual may change sexes during its lifetime.
It might seem that since only some of the male descendants of Queen Victoria suffered from hemophilia, the Y chromosome was the culprit. However, the pattern of inheritance was through the female line of descent, which meant it had to be the X chromosome, and it turned out that in fact the genetic defect that causes hemophilia actually is on the X chromosome.
The reason that females don't suffer from hemophilia is because the "good" gene on the other X chromosome continues to operate correctly, producing a working blood-clotting system, and compensating for the disabled or "bad" gene on the other X chromosome. The Y chromosome of the male doesn't carry the critical genes for the blood-clotting system at all, and so his blood-clotting system is disabled. Since the females carrying the "bad" gene have two X chromosomes, one "bad" and one "good", the odds of her male children suffering from hemophilia are fifty-fifty. That was what was observed in practice for the descendants of Queen Victoria.
As this explanation shows, it is actually not impossible for a female to be a hemophiliac. If both X chromosomes of a female are "bad", she will suffer from the syndrome. However, this would require a fair amount of inbreeding, since the hemophilia gene is rare in the general population -- of course it is, since the selection against it is severe, its effect is so disastrous that sooner or later it kills off the males carrying it. Males who suffer from hemophilia have much less chance to reproduce than healthy males, and their affliction is apparent anyway, meaning any female who selects one as a mate knows she's taking chances with her children, whether she has any technical understanding of heredity or not.
Hemophilia was not known in Queen Victoria's ancestors. The unproveable suspicion is that the mutation arose in her father, the Duke of Kent. Human females acquire a stockpile of ova at sexual maturity, with the ova then doled out over the rest of their reproductive lifetime. Male humans, in contrast, produce sperm over their entire reproductive lifetime, giving many more possibilities for accidents to happen, for mutations to arise. Incidentally, although Prince Charles can trace his line of ancestry back to Queen Victoria through both of his parents, Queen Elizabeth II and Prince Philip, neither of those lines carried the bogus X chromosome that causes hemophilia.
* As this story suggests, the action of chromosomes can be complicated. It's worth noting here as a footnote for later that in some cases very different species can sometimes be hybridized by one or both parents contributing a dual set, not as normal a single set, of chromosomes to its progeny, a scheme known as "polyploidy". It is much more common in plants than it is in animals; modern wheat, for example, is "hexaploid", with three sets of chromosomes, or six chromosomes in all.
In any case, Morgan had finally put genetics on a sound scientific basis, establishing it is as a matter of "hard heredity". Morgan remains a prophet of great honor, his work opening doors that up to that time had seemed securely locked.
BACK_TO_TOP* Another weakness of Darwinian evolutionary theory was the lack of information on human predecessors, but by the early 20th century that issue was gradually being addressed as well. The pioneer was a Dutch biologist named Eugene Dubois (1858:1940) who had been much impressed by Darwinian thinking in his youth. He began his career as a professor of anatomy at the University of Amsterdam, working on the comparative morphology of humans and animals. He soon set his sights on more ambitious research.
Darwin had suggested in DESCENT OF MAN that humans had arisen in Africa, but Lyell had observed in ANTIQUITY OF MAN that the most humanlike of the apes, the orangutan, the "Old Man Of The Woods" as locals called him, was from Southeast Asia, suggesting an alternate origin of humankind. Haeckel also favored an Asian origin of humankind, though his reasoning was based more on racial prejudice than evidence. Dubois favored Asian origins as well, and since the East Indies were part of the Dutch colonial empire at the time, he had a good place to start looking. He left the University of Amsterdam in 1887 and signed on as a medical doctor with the Dutch colonial army.
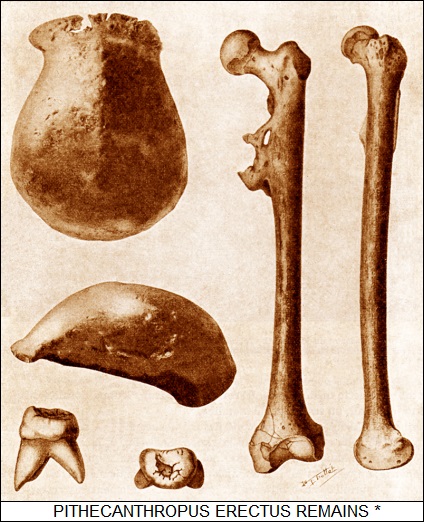
On arriving in the East Indies, Dubois spent four years hunting for prehuman remains on the islands of Java and Sumatra. The whole idea might have seemed crankish and foolish to Dubois' scientific contemporaries -- except for the fact that he succeeded. In 1891, while digging in Java, his team found a fossil molar of some species of human and a skullcap that suggested the brain case of this particular species was substantially smaller than that of a modern human. A year later, in 1892, a thigh bone was discovered that suggested the species walked upright.
Dubois named the find Pithecanthropus erectus or "Upright Apeman", though it more popularly became known as "Java Man". Dubois claimed it was the "missing link" between apes and humans. Given the sketchiness of the find -- a molar, skullcap, and thighbone wasn't much to go on -- not all of his colleagues were impressed, some claiming the thing was just another ape and others wondering if all three fossil samples were even from the same creature. Dubois, who had returned to the Netherlands in 1895, became snarled up in the controversy, becoming more defensive to the point of outright paranoia, unwilling to let go of his samples for examination.
To confuse matters, in 1912 what appeared to be a fossil human was found at Piltdown in England, with a humanlike upper skull and an apelike jaw. Many were suspicious since the jaw joint elements were missing, suggesting that the "Piltdown Man" was a hoax, a plant. It would take over 40 years to learn that the Piltdown Man "fossil" was a human skull with an orangutan jaw. Nobody would ever know for certain who set up the hoax or why, but it did much to muddy the waters for years -- at least, in an era where the sketchiness of data made the waters easy to muddy.
Despite the obstacles, events began to lend weight to Dubois' claims. More fossils of a similar creature were found in China from 1929 and named "Peking Man". In time there was no doubt that these fossils were of an extinct humanlike species, which became known as Homo erectus; however, they were too similar to humans to really be much of a "missing link" with apes.
* By that time, other candidates were available. In 1924, a South African student found a fossil skull at a limestone quarry near the town of Taung. She handed the skull over to Raymond Dart (1893:1988), her Australian-born anatomy professor, who judged it to be a baboon skull at first. He requested that the quarry owner kindly send him any more fossils found there, and the owner obliged, sending two crates of neatly-packed fossils in the fall of 1924.
The fossils included enough of a skull of a child to permit a good reconstruction. He immediately recognized that the Taung fossils were not those of baboons; he excitedly announced in the scientific press in early 1925 that his discovery, which he called Australopithecus africanus or "Southern Ape Of Africa", labeling it "an extinct link between man and his simian ancestor." Dart's notions were not universally popular among his scientific colleagues, in part because he was proposing that the savannas of Africa and not the jungles of Asia were the "cradle of humanity".
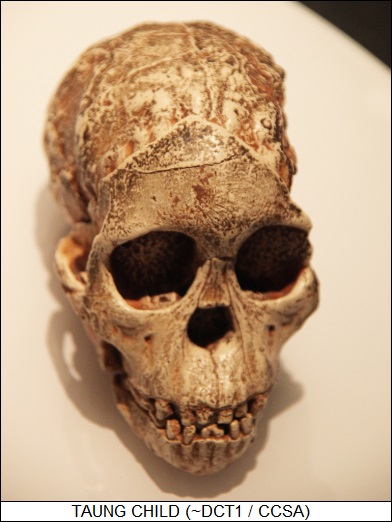
Dart did have one enthusiastic advocate, a Scots-born anthropologist named Robert Broom (1866:1951), a recognized expert in African fossils. He was impressed by Dart's "Taung child" and accepted Dart's conclusions, conducting expeditions to hunt for more fossils -- achieving success by finding remains of a half-dozen "australopithecines" at Sterkfontein, South Africa, in 1936. Broom also discovered fossil remains of a "heavy" species of australopithecine, which would eventually become known as Australopithecus robustus. It was becoming apparent that Dart had been on the right track, though the fossil evidence still remained sketchy and there was plenty of room for argument. Further progress was sidetracked for the moment by World War II: people had other things to worry about.
* Incidentally, while the term "missing link" remains a pop-science cliche, researchers tend to dislike the term, not only because it is overused, but because it is misleading. The idea of a "missing link" implies some nice neat sequence of species in an evolutionary chain -- species A leading to species B leading to species C -- but that's not how evolution really works. It's species branching off from each other repeatedly, much more like a tree than a chain.
The term "intermediate form" is preferred. It simply means that one species has features between two other species -- that is, species B has some features of earlier species A and some features of later species C. This usage recognizes the emergence of features along the evolutionary tree of a group of organisms, but avoids the implication that there is a direct ancestral line between the three species. It may not be true that species A was the "parent" of species B, and species B was the "parent" of species C; species A may have instead been an "uncle" in some degree of species B, and species B may have an "uncle" in turn of species C. The reality remains of a succession of earlier forms along the path to a modern species.
BACK_TO_TOP